

The mutualism-antagonism continuum in Neotropical palm-frugivore interactions: from interaction outcomes to ecosystem dynamics
- PMID: 34725900
- PMCID: PMC9297963
- DOI: 10.1111/brv.12809
The mutualism-antagonism continuum in Neotropical palm-frugivore interactions: from interaction outcomes to ecosystem dynamics
Abstract
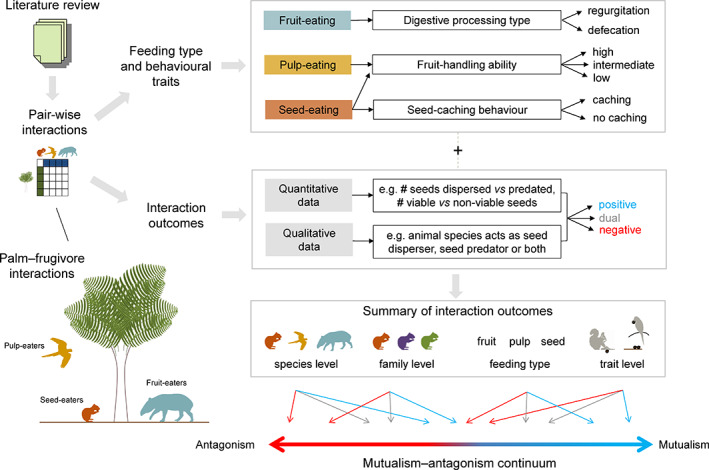
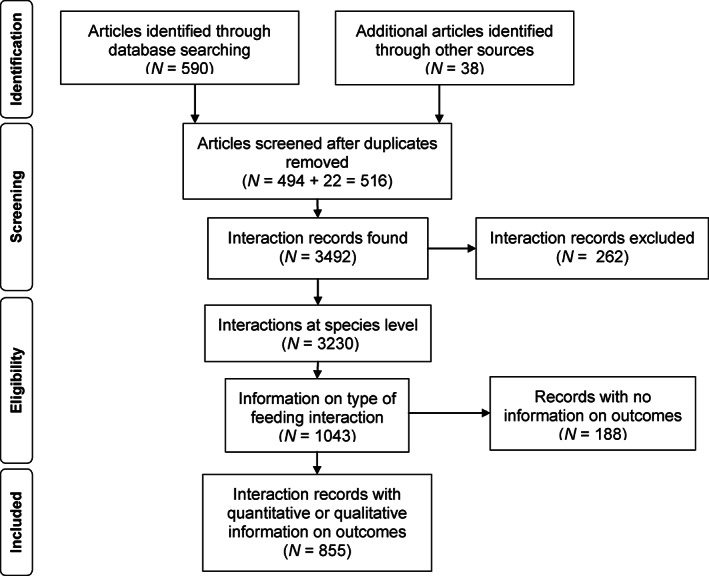
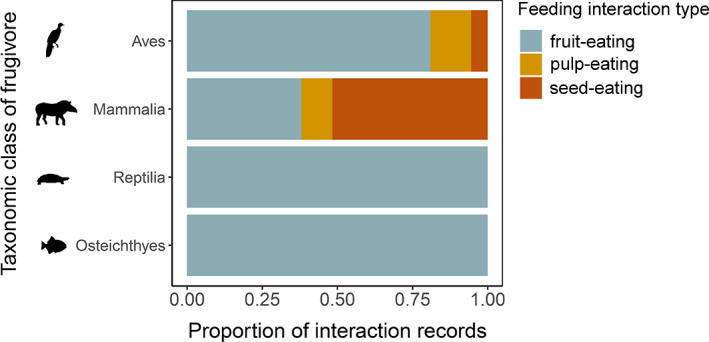
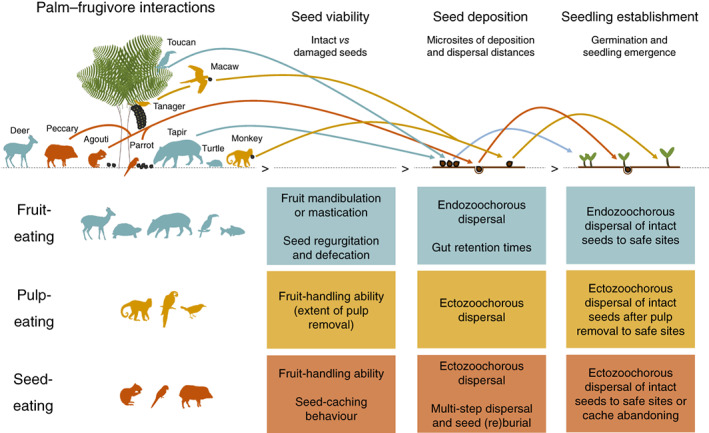
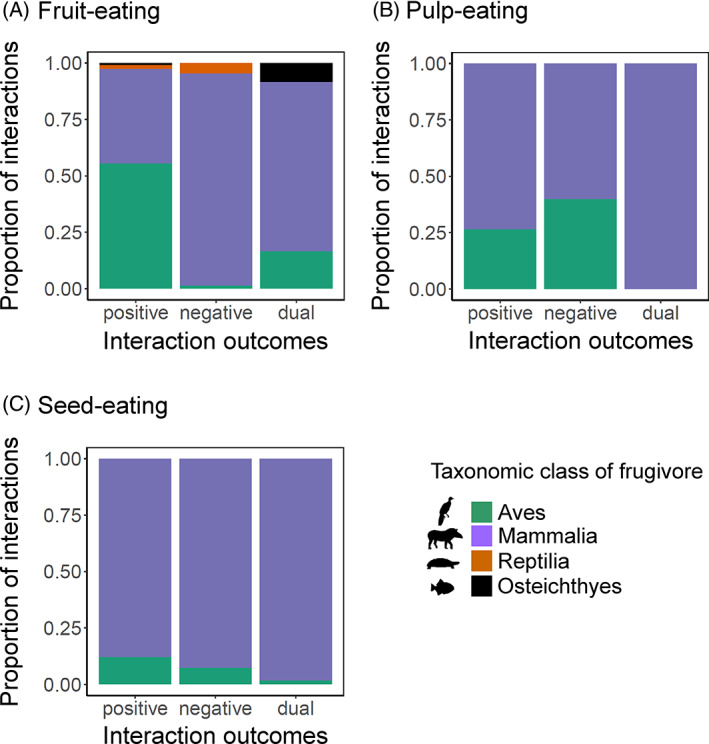
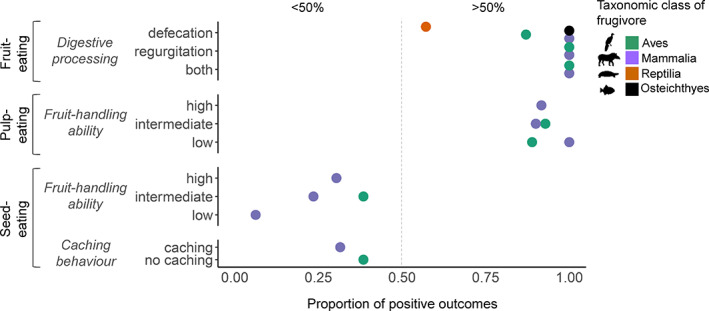
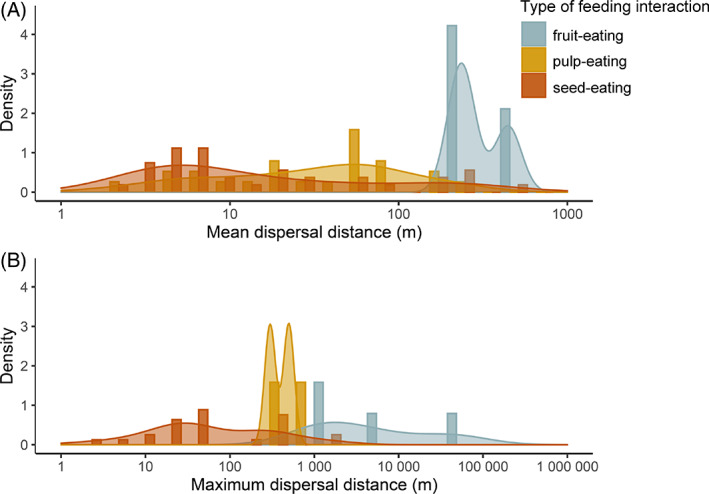
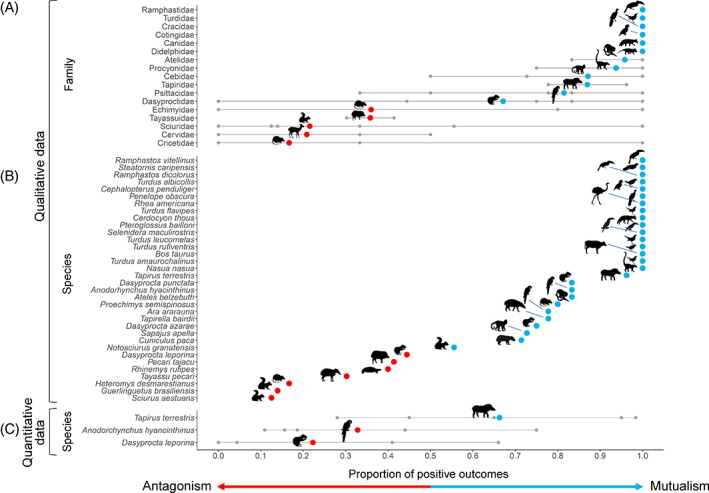
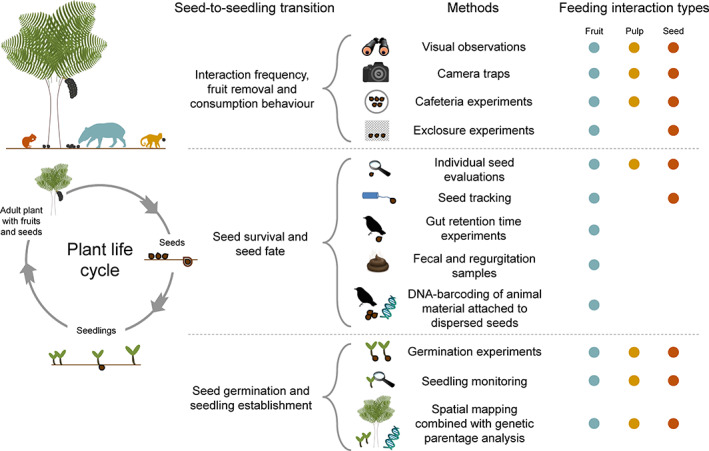
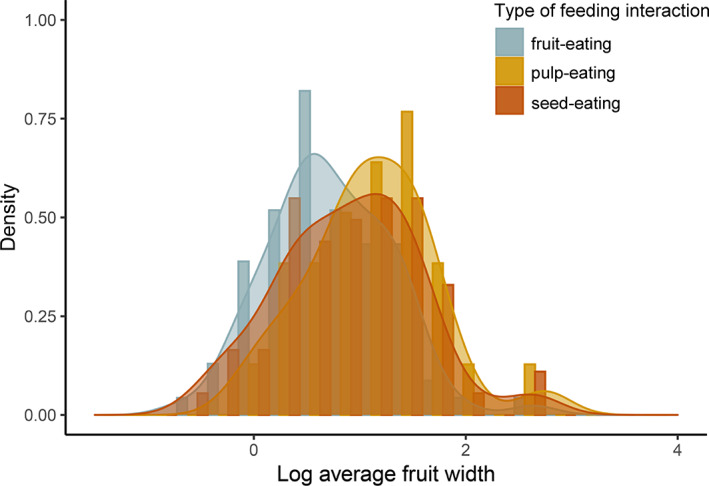
Frugivory, that is feeding on fruits, pulp or seeds by animals, is usually considered a mutualism when interactions involve seed dispersal, and an antagonism when it results in the predation and destruction of seeds. Nevertheless, most frugivory interactions involve both benefits and disadvantages for plants, and the net interaction outcomes thus tend to vary along a continuum from mutualism to antagonism. Quantifying outcome variation is challenging and the ecological contribution of frugivorous animals to plant demography thus remains little explored. This is particularly true for interactions in which animals do not ingest entire fruits, that is in seed-eating and pulp-eating. Here, we provide a comprehensive review of Neotropical palm-frugivore interactions, with a focus on how frugivore consumption behaviour (i.e. digestive processing, fruit-handling ability and caching behaviour) and feeding types (fruit-eating, pulp-eating and seed-eating) influence interaction outcomes at different demographic stages of palms. We compiled a total of 1043 species-level palm-frugivore interaction records that explicitly captured information on which parts of palm fruits are eaten by animals. These records showed consumption of fruits of 106 Neotropical palm species by 273 vertebrate species, especially birds (50%) and mammals (45%), but also fish (3%) and reptiles (2%). Fruit-eating involved all four taxonomic vertebrate classes whereas seed-eating and pulp-eating were only recorded among birds and mammals. Most fruit-eating interactions (77%) resulted in positive interaction outcomes for plants (e.g. gut-passed seeds are viable or seeds are successfully dispersed), regardless of the digestive processing type of vertebrate consumers (seed defecation versus regurgitation). The majority of pulp-eating interactions (91%) also resulted in positive interaction outcomes, for instance via pulp removal that promoted seed germination or via dispersal of intact palm seeds by external transport, especially if animals have a good fruit-handling ability (e.g. primates, and some parrots). By contrast, seed-eating interactions mostly resulted in dual outcomes (60%), where interactions had both negative effects on seed survival and positive outcomes through seed caching and external (non-digestive) seed dispersal. A detailed synthesis of available field studies with qualitative and quantitative information provided evidence that 12 families and 27 species of mammals and birds are predominantly on the mutualistic side of the continuum whereas five mammalian families, six mammal and one reptile species are on the antagonistic side. The synthesis also revealed that most species can act as partial mutualists, even if they are typically considered antagonists. Our review demonstrates how different consumption behaviours and feeding types of vertebrate fruit consumers can influence seed dispersal and regeneration of palms, and thus ultimately affect the structure and functioning of tropical ecosystems. Variation in feeding types of animal consumers will influence ecosystem dynamics via effects on plant population dynamics and differences in long-distance seed dispersal, and may subsequently affect ecosystem functions such as carbon storage. The quantification of intra- and inter-specific variation in outcomes of plant-frugivore interactions - and their positive and negative effects on the seed-to-seedling transition of animal-dispersed plants - should be a key research focus to understand better the mutualism-antagonism continuum and its importance for ecosystem dynamics.
Keywords: Arecaceae; ecosystem functioning; ectozoochory; endozoochory; granivory; palm endocarp; seed caching; seed dispersal distance; seed predation; vertebrate.
© 2021 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures

Similar articles
-
Moving from frugivory to seed dispersal: Incorporating the functional outcomes of interactions in plant-frugivore networks.J Anim Ecol. 2018 Jul;87(4):995-1007. doi: 10.1111/1365-2656.12831. Epub 2018 Apr 20. J Anim Ecol. 2018. PMID: 29603211 Free PMC article.
-
Frugivory and seed dispersal in a hyperdiverse plant clade and its role as a keystone resource for the Neotropical fauna.Ann Bot. 2021 Apr 17;127(5):577-595. doi: 10.1093/aob/mcaa189. Ann Bot. 2021. PMID: 33151331 Free PMC article.
-
Seed dispersal as an ecosystem service: frugivore loss leads to decline of a socially valued plant, Capsicum frutescens.Ecol Appl. 2018 Apr;28(3):655-667. doi: 10.1002/eap.1667. Epub 2018 Feb 27. Ecol Appl. 2018. PMID: 29271019 Free PMC article.
-
Neotropical fish-fruit interactions: eco-evolutionary dynamics and conservation.Biol Rev Camb Philos Soc. 2015 Nov;90(4):1263-78. doi: 10.1111/brv.12153. Epub 2015 Jan 19. Biol Rev Camb Philos Soc. 2015. PMID: 25599800 Review.
-
Frugivory and seed dispersal in the Galápagos: what is the state of the art?Integr Zool. 2011 Jun;6(2):110-129. doi: 10.1111/j.1749-4877.2011.00236.x. Integr Zool. 2011. PMID: 21645276 Review.
Cited by
-
Novel plant-frugivore network on Mauritius is unlikely to compensate for the extinction of seed dispersers.Nat Commun. 2023 Feb 23;14(1):1019. doi: 10.1038/s41467-023-36669-9. Nat Commun. 2023. PMID: 36823195 Free PMC article.
-
The megaherbivore gap after the non-avian dinosaur extinctions modified trait evolution and diversification of tropical palms.Proc Biol Sci. 2022 Apr 13;289(1972):20212633. doi: 10.1098/rspb.2021.2633. Epub 2022 Apr 13. Proc Biol Sci. 2022. PMID: 35414237 Free PMC article.
-
Precipitation Drives Frugivory in a Subtropical Generalist Herbivore, the Gopher Tortoise, and Alters Its Functional Role as a Seed Disperser.Ecol Evol. 2024 Nov 18;14(11):e70585. doi: 10.1002/ece3.70585. eCollection 2024 Nov. Ecol Evol. 2024. PMID: 39559471 Free PMC article.
-
From friend to foe and back: coevolutionary transitions in the mutualism-antagonism continuum.Proc Biol Sci. 2025 Jul;292(2051):20242326. doi: 10.1098/rspb.2024.2326. Epub 2025 Jul 23. Proc Biol Sci. 2025. PMID: 40695341 Free PMC article.
-
What drives seed dispersal effectiveness?Ecol Evol. 2023 Aug 31;13(9):e10459. doi: 10.1002/ece3.10459. eCollection 2023 Sep. Ecol Evol. 2023. PMID: 37664501 Free PMC article.
References
-
- Acevedo‐Quintero, J. F. & Zamora‐Abrego, J. G. (2016). Role of mammals on seed dispersal and predation processes of Mauritia flexuosa (Arecaceae) in the Colombian Amazon. Revista de Biologia Tropical 64, 5–15. - PubMed
-
- Acevedo‐Quintero, J. F. , Zamora‐Abrego, J. G. & Ortega‐León, Á. M. (2018). The prickles of Astrocaryum malybo as a structural defense to avoid seed predation. Food Webs 16, e00088.
-
- * Adler, G. H. & Kestell, D. W. (1998). Fates of Neotropical tree seeds influenced by spiny rats (Proechimys semispinosus). Biotropica 30, 677–681.
-
- * Aliaga‐Rossel, E. & Fragoso, J. M. (2015). Defaunation affects Astrocaryum gratum (Arecales: Arecaceae) seed survivorship in a sub‐montane tropical forest. Revista de Biología Tropical 63, 57–67. - PubMed
-
- * Alves, B. C. , Mendes, C. P. & Ribeiro, M. C. (2018). Queen palm fruit selection and foraging techniques of squirrels in the Atlantic Forest. Biotropica 50, 274–281.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
