

ERAP1, ERAP2, and Two Copies of HLA-Aw19 Alleles Increase the Risk for Birdshot Chorioretinopathy in HLA-A29 Carriers
- PMID: 34727153
- PMCID: PMC8572510
- DOI: 10.1167/iovs.62.14.3
ERAP1, ERAP2, and Two Copies of HLA-Aw19 Alleles Increase the Risk for Birdshot Chorioretinopathy in HLA-A29 Carriers
Abstract
Purpose: Birdshot chorioretinopathy (BSCR) is strongly associated with HLA-A29. This study was designed to elucidate the genetic modifiers of BSCR in HLA-A29 carriers.
Methods: We sequenced the largest BSCR cohort to date, including 286 cases and 108 HLA-A29-positive controls to determine genome-wide common and rare variant associations. We further typed the HLA alleles of cases and 45,386 HLA-A29 controls of European ancestry to identify HLA alleles that associate with BSCR risk.
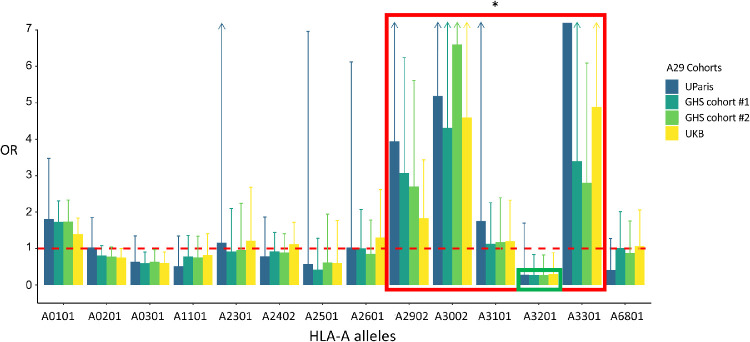
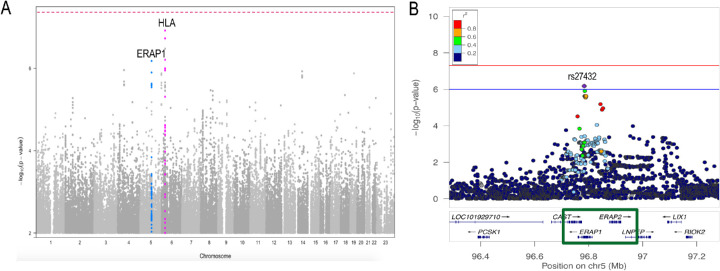
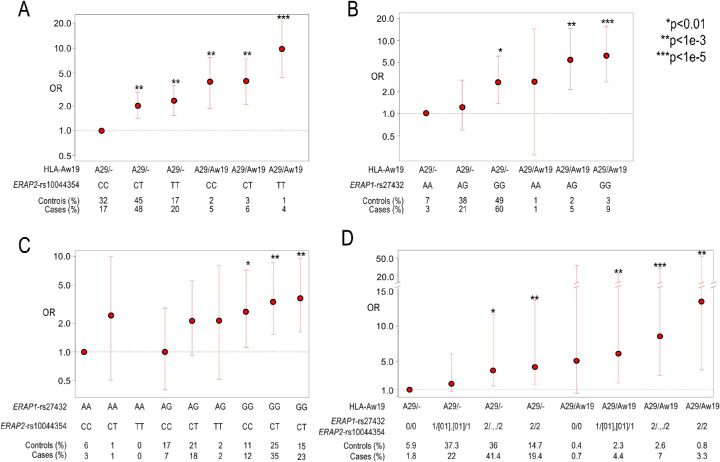
Results: Carrying a second allele that belongs to the HLA-Aw19 broad antigen family (including HLA-A29, -A30, -A31, and -A33) increases the risk for BSCR (odds ratio [OR] = 4.44; P = 2.2e-03). This result was validated by comparing allele frequencies to large HLA-A29-controlled cohorts (n = 45,386; OR > 2.5; P < 1.3e-06). We also confirm that ERAP1 and ERAP2 haplotypes modulate disease risk. A meta-analysis with an independent dataset confirmed that ERAP1 and ERAP2 haplotypes modulate the risk for disease at a genome-wide significant level: ERAP1-rs27432 (OR = 2.46; 95% confidence interval [CI], 1.85-3.26; P = 4.07e-10), an expression quantitative trait locus (eQTL) decreasing ERAP1 expression; and ERAP2-rs10044354 (OR = 1.95; 95% CI, 1.55-2.44; P = 6.2e-09), an eQTL increasing ERAP2 expression. Furthermore, ERAP2-rs2248374 that disrupts ERAP2 expression is protective (OR = 0.56; 95% CI, 0.45-0.70; P = 2.39e-07). BSCR risk is additively increased when combining ERAP1/ERAP2 risk genotypes with two copies of HLA-Aw19 alleles (OR = 13.53; 95% CI, 3.79-54.77; P = 1.17e-05).
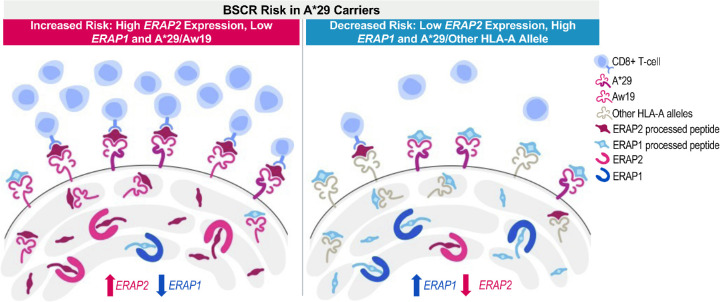
Conclusions: The genetic factors increasing BSCR risk demonstrate a pattern of increased processing, as well as increased presentation of ERAP2-specific peptides. This suggests a mechanism in which exceeding a peptide presentation threshold activates the immune response in choroids of A29 carriers.
Conflict of interest statement
Disclosure:
Figures
References
-
- Shao EH, Menezo V, Taylor SR. Birdshot chorioretinopathy. Curr Opin Ophthalmol. 2014; 25: 488–494. - PubMed
-
- Ryan SJ, Maumenee AE. Birdshot retinochoroidopathy. Am J Ophthalmol. 1980; 89: 31–45. - PubMed
-
- Nussenblatt RB, Mittal KK, Ryan S, Green WR, Maumenee AE. Birdshot retinochoroidopathy associated with HLA-A29 antigen and immune responsiveness to retinal S-antigen. Am J Ophthalmol. 1982; 94: 147–158. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
