

Coupling plant litter quantity to a novel metric for litter quality explains C storage changes in a thawing permafrost peatland
- PMID: 34727401
- PMCID: PMC9298822
- DOI: 10.1111/gcb.15970
Coupling plant litter quantity to a novel metric for litter quality explains C storage changes in a thawing permafrost peatland
Abstract
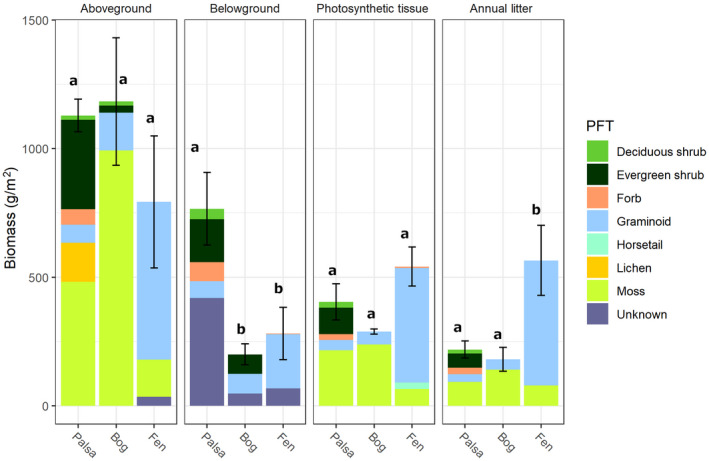
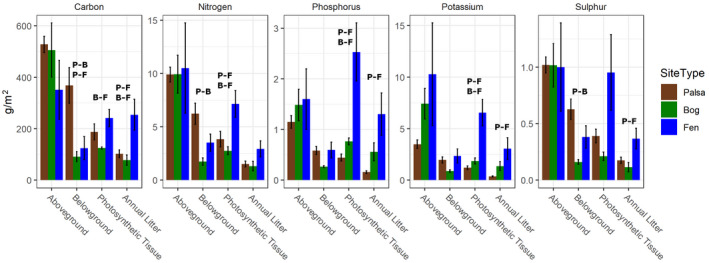
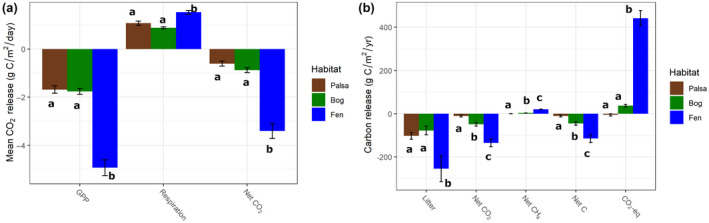
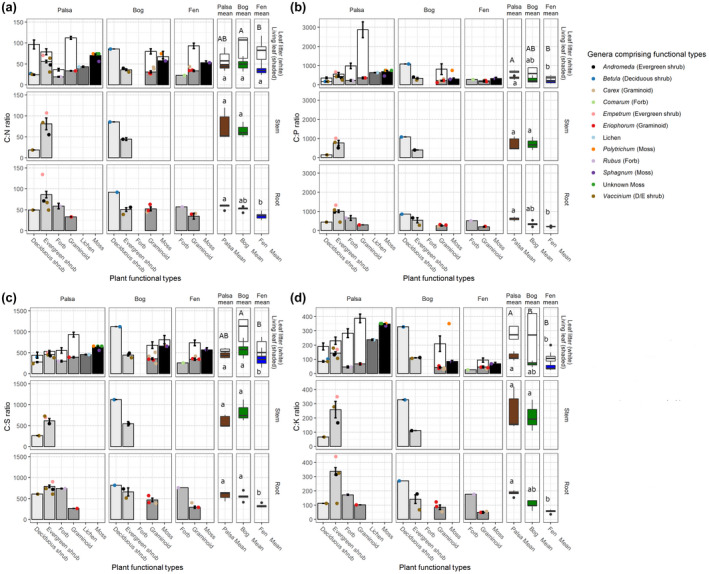
Permafrost thaw is a major potential feedback source to climate change as it can drive the increased release of greenhouse gases carbon dioxide (CO2 ) and methane (CH4 ). This carbon release from the decomposition of thawing soil organic material can be mitigated by increased net primary productivity (NPP) caused by warming, increasing atmospheric CO2 , and plant community transition. However, the net effect on C storage also depends on how these plant community changes alter plant litter quantity, quality, and decomposition rates. Predicting decomposition rates based on litter quality remains challenging, but a promising new way forward is to incorporate measures of the energetic favorability to soil microbes of plant biomass decomposition. We asked how the variation in one such measure, the nominal oxidation state of carbon (NOSC), interacts with changing quantities of plant material inputs to influence the net C balance of a thawing permafrost peatland. We found: (1) Plant productivity (NPP) increased post-thaw, but instead of contributing to increased standing biomass, it increased plant biomass turnover via increased litter inputs to soil; (2) Plant litter thermodynamic favorability (NOSC) and decomposition rate both increased post-thaw, despite limited changes in bulk C:N ratios; (3) these increases caused the higher NPP to cycle more rapidly through both plants and soil, contributing to higher CO2 and CH4 fluxes from decomposition. Thus, the increased C-storage expected from higher productivity was limited and the high global warming potential of CH4 contributed a net positive warming effect. Although post-thaw peatlands are currently C sinks due to high NPP offsetting high CO2 release, this status is very sensitive to the plant community's litter input rate and quality. Integration of novel bioavailability metrics based on litter chemistry, including NOSC, into studies of ecosystem dynamics, is needed to improve the understanding of controls on arctic C stocks under continued ecosystem transition.
Keywords: C storage; NOSC; Stordalen Mire; decomposition; litter chemistry; peat; permafrost thaw; plant community change.
© 2021 The Authors. Global Change Biology published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
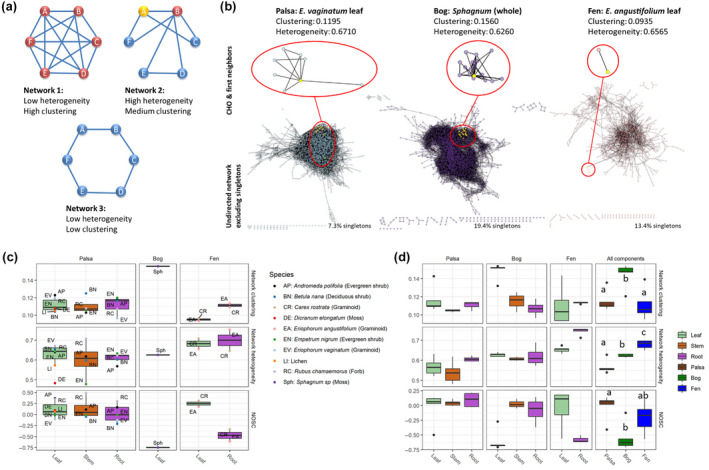
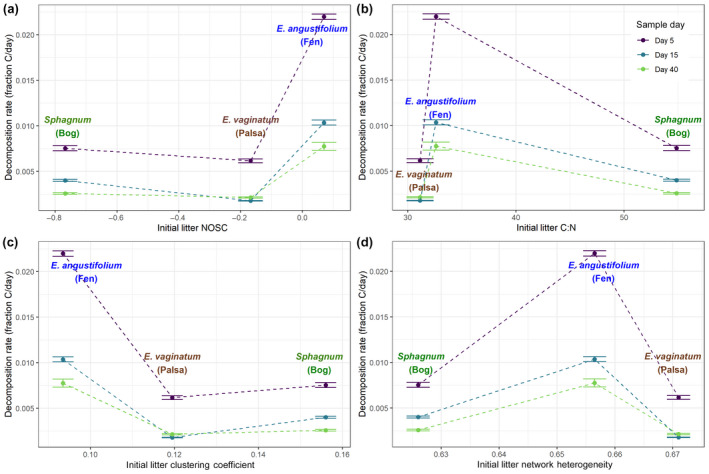
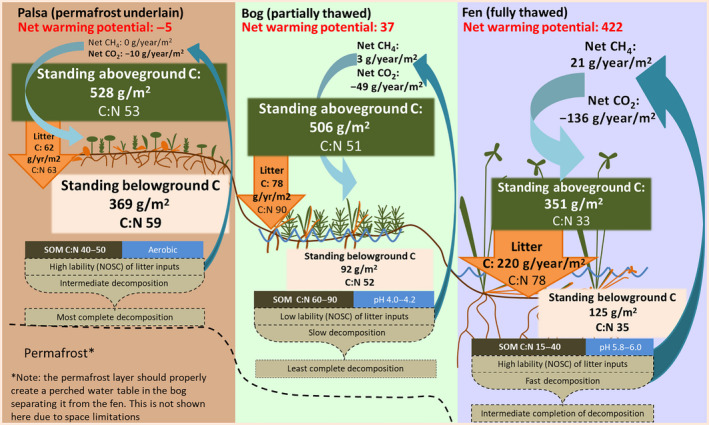
References
-
- Aerts, R. (1996). nutrient resorption from senescing leaves of perennials: Are there general patterns? The Journal of Ecology, 84(4), 597–608. 10.2307/2261481 - DOI
-
- AminiTabrizi, R. , Wilson, R. M. , Fudyma, J. D. , Hodgkins, S. B. , Heyman, H. M. , Rich, V. I. , Saleska, S. R. , Chanton, J. P. , & Tfaily, M. M. (2020). Controls on soil organic matter degradation and subsequent greenhouse gas emissions across a permafrost thaw gradient in Northern Sweden. Frontiers in Earth. 10.3389/feart.2020.557961 - DOI
-
- Bäckstrand, K. , Crill, P. M. , Jackowicz‐Korczyñski, M. , Mastepanov, M. , Christensen, T. R. , & Bastviken, D. (2010). Annual carbon gas budget for a subarctic peatland, Northern Sweden. Biogeosciences, 7(1), 95–108. 10.5194/bg-7-95-2010 - DOI
-
- Bintanja, R. , & Andry, O. (2017). Towards a rain‐dominated Arctic. Nature Climate Change, 7(4), 263–267. 10.1038/nclimate3240 - DOI
