

Cellular Plasticity, Reprogramming, and Regeneration: Metaplasia in the Stomach and Beyond
- PMID: 34728185
- PMCID: PMC8792220
- DOI: 10.1053/j.gastro.2021.10.036
Cellular Plasticity, Reprogramming, and Regeneration: Metaplasia in the Stomach and Beyond
Abstract
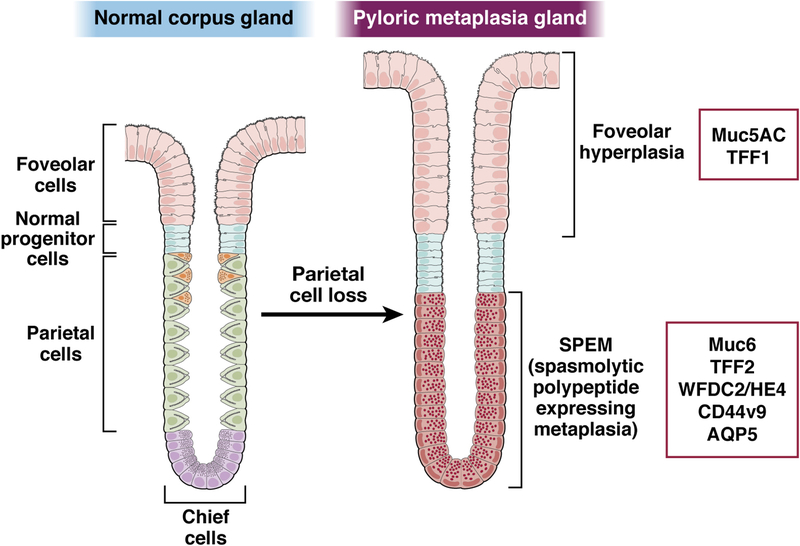
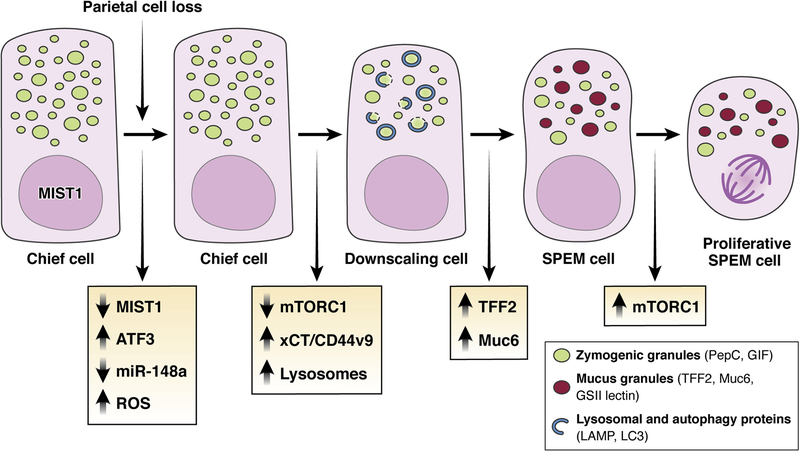
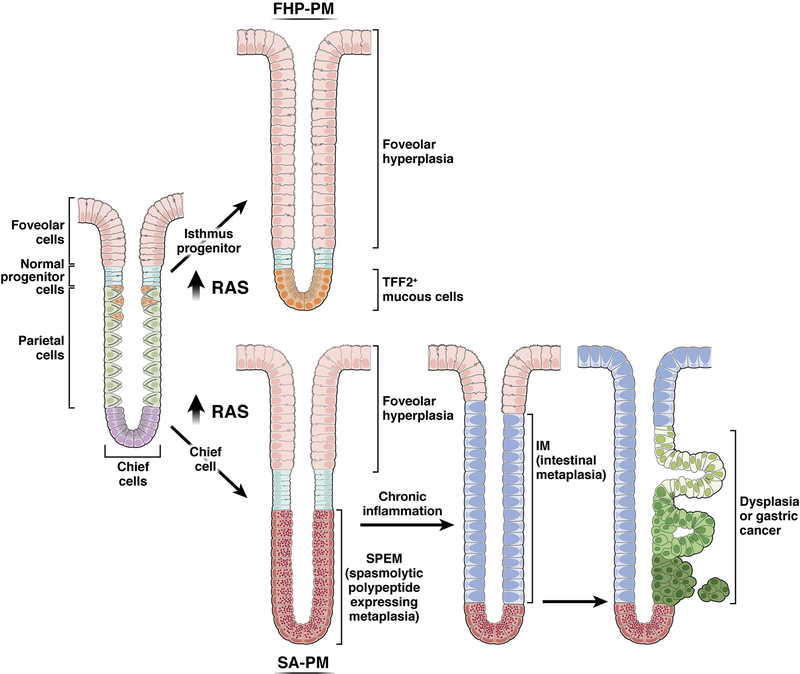
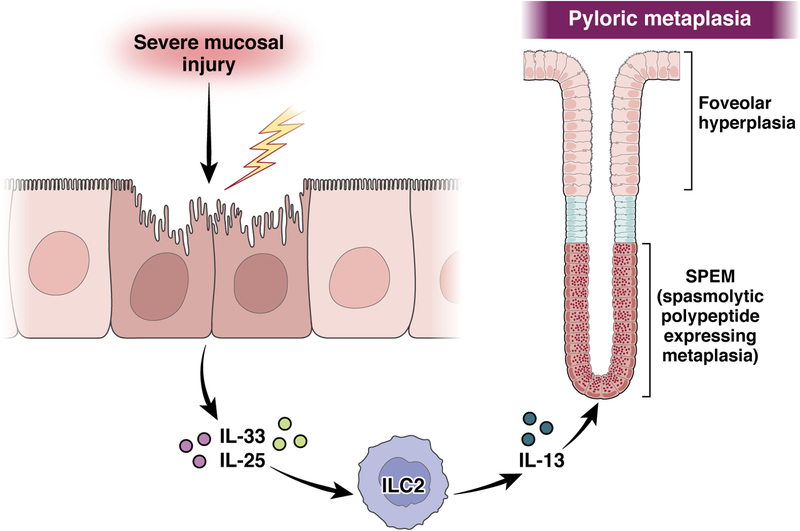
The mucosa of the body of the stomach (ie, the gastric corpus) uses 2 overlapping, depth-dependent mechanisms to respond to injury. Superficial injury heals via surface cells with histopathologic changes like foveolar hyperplasia. Deeper, usually chronic, injury/inflammation, most frequently induced by the carcinogenic bacteria Helicobacter pylori, elicits glandular histopathologic alterations, initially manifesting as pyloric (also known as pseudopyloric) metaplasia. In this pyloric metaplasia, corpus glands become antrum (pylorus)-like with loss of acid-secreting parietal cells (atrophic gastritis), expansion of foveolar cells, and reprogramming of digestive enzyme-secreting chief cells into deep antral gland-like mucous cells. After acute parietal cell loss, chief cells can reprogram through an orderly stepwise progression (paligenosis) initiated by interleukin-13-secreting innate lymphoid cells (ILC2s). First, massive lysosomal activation helps mitigate reactive oxygen species and remove damaged organelles. Second, mucus and wound-healing proteins (eg, TFF2) and other transcriptional alterations are induced, at which point the reprogrammed chief cells are recognized as mucus-secreting spasmolytic polypeptide-expressing metaplasia cells. In chronic severe injury, glands with pyloric metaplasia can harbor both actively proliferating spasmolytic polypeptide-expressing metaplasia cells and eventually intestine-like cells. Gastric glands with such lineage confusion (mixed incomplete intestinal metaplasia and proliferative spasmolytic polypeptide-expressing metaplasia) may be at particular risk for progression to dysplasia and cancer. A pyloric-like pattern of metaplasia after injury also occurs in other gastrointestinal organs including esophagus, pancreas, and intestines, and the paligenosis program itself seems broadly conserved across tissues and species. Here we discuss aspects of metaplasia in stomach, incorporating data derived from animal models and work on human cells and tissues in correlation with diagnostic and clinical implications.
Copyright © 2022 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no conflicts of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
- 27145/CRUK_/Cancer Research UK/United Kingdom
- R01 DK101332/DK/NIDDK NIH HHS/United States
- I01 BX000930/BX/BLRD VA/United States
- P30 DK056338/DK/NIDDK NIH HHS/United States
- R01 CA246208/CA/NCI NIH HHS/United States
- R01 DK120680/DK/NIDDK NIH HHS/United States
- R01 DK110406/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- U54 CA163060/CA/NCI NIH HHS/United States
- R01 DK105129/DK/NIDDK NIH HHS/United States
- R01 CA239645/CA/NCI NIH HHS/United States
- R01 DK094989/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
