

Hypoxia induces stress fiber formation in adipocytes in the early stage of obesity
- PMID: 34728615
- PMCID: PMC8563745
- DOI: 10.1038/s41598-021-00335-1
Hypoxia induces stress fiber formation in adipocytes in the early stage of obesity
Abstract
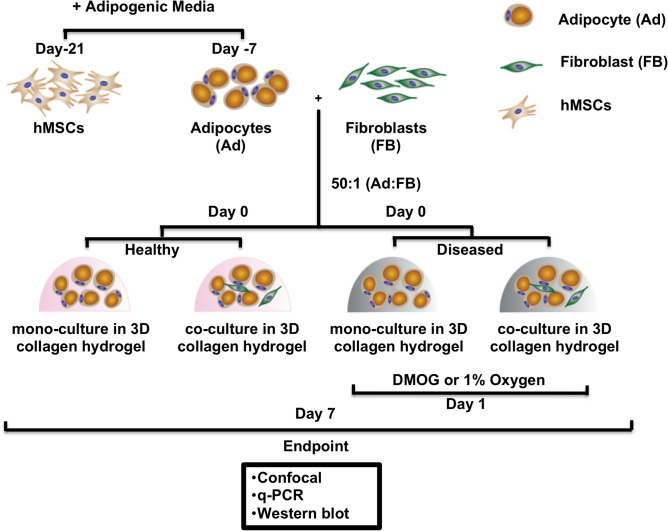
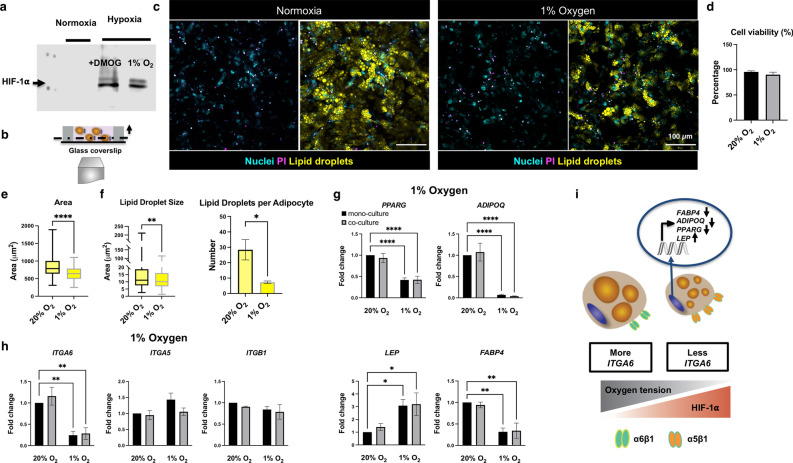
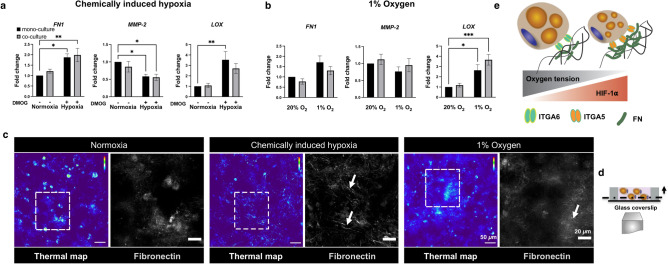
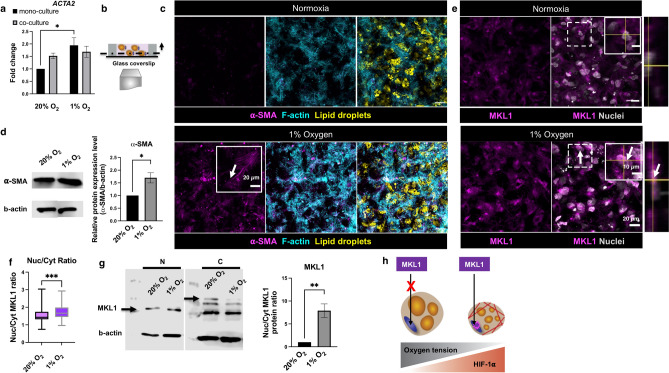
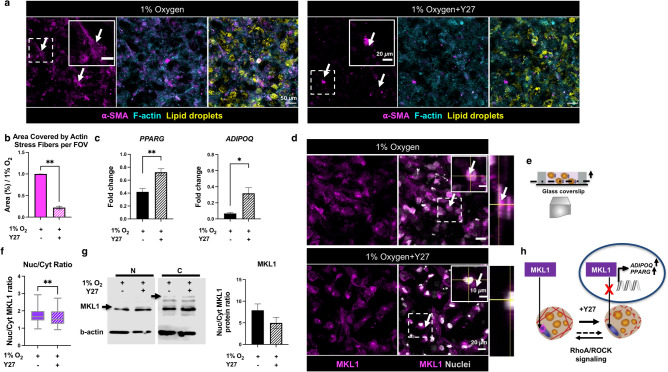
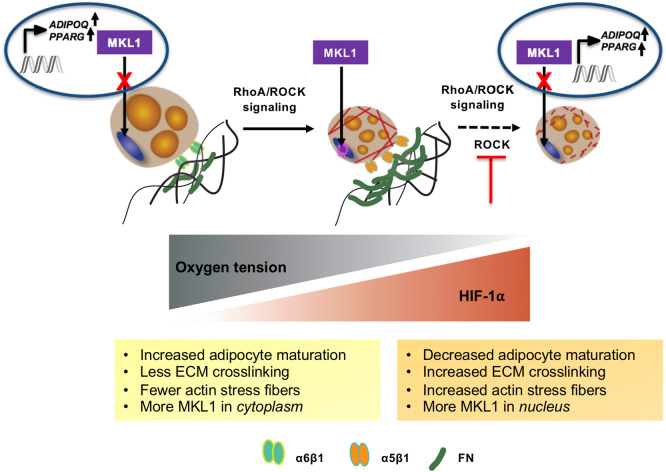
In obese adipose tissue (AT), hypertrophic expansion of adipocytes is not matched by new vessel formation, leading to AT hypoxia. As a result, hypoxia inducible factor-1⍺ (HIF-1⍺) accumulates in adipocytes inducing a transcriptional program that upregulates profibrotic genes and biosynthetic enzymes such as lysyl oxidase (LOX) synthesis. This excess synthesis and crosslinking of extracellular matrix (ECM) components cause AT fibrosis. Although fibrosis is a hallmark of obese AT, the role of fibroblasts, cells known to regulate fibrosis in other fibrosis-prone tissues, is not well studied. Here we have developed an in vitro model of AT to study adipocyte-fibroblast crosstalk in a hypoxic environment. Further, this in vitro model was used to investigate the effect of hypoxia on adipocyte mechanical properties via ras homolog gene family member A (RhoA)/Rho-associated coiled-coil kinases (ROCK) signaling pathways. We confirmed that hypoxia creates a diseased phenotype by inhibiting adipocyte maturation and inducing actin stress fiber formation facilitated by myocardin-related transcription factor A (MRTF-A/MKL1) nuclear translocation. This work presents new potential therapeutic targets for obesity by improving adipocyte maturation and limiting mechanical stress in obese AT.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
