

Bacterial lectin BambL acts as a B cell superantigen
- PMID: 34731252
- PMCID: PMC8629787
- DOI: 10.1007/s00018-021-04009-z
Bacterial lectin BambL acts as a B cell superantigen
Abstract
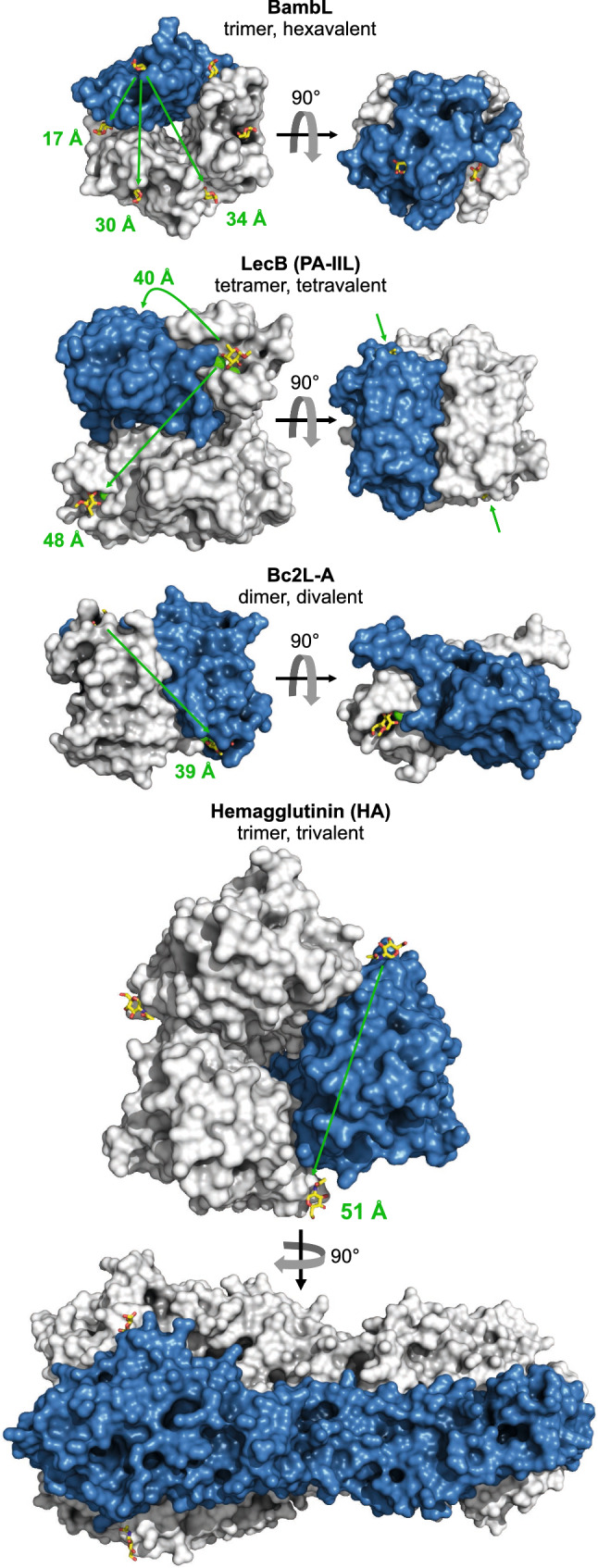
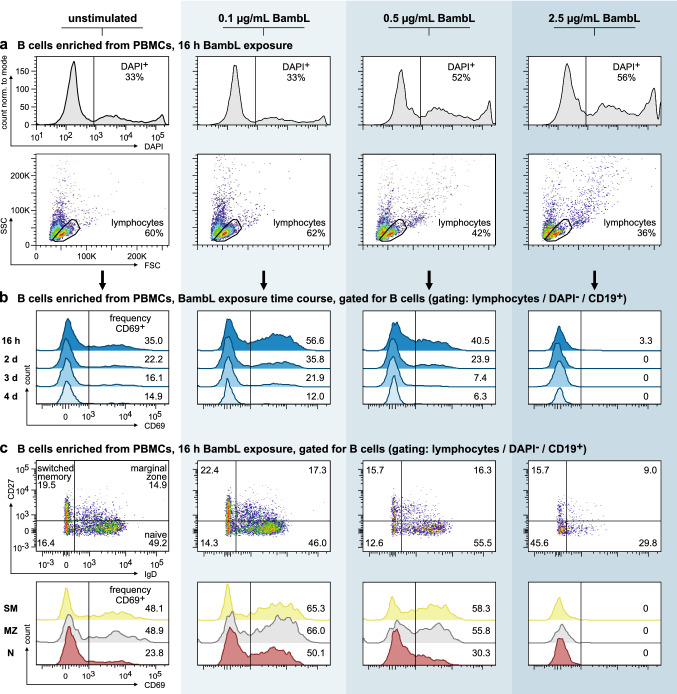
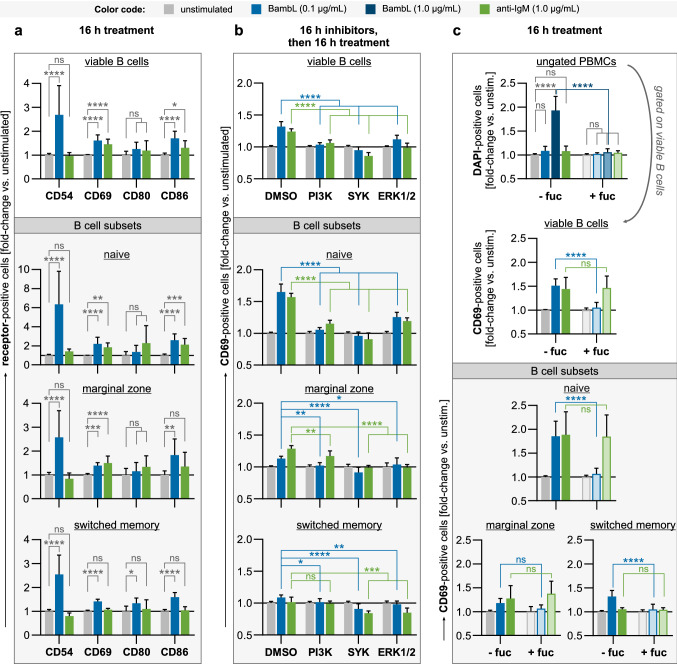
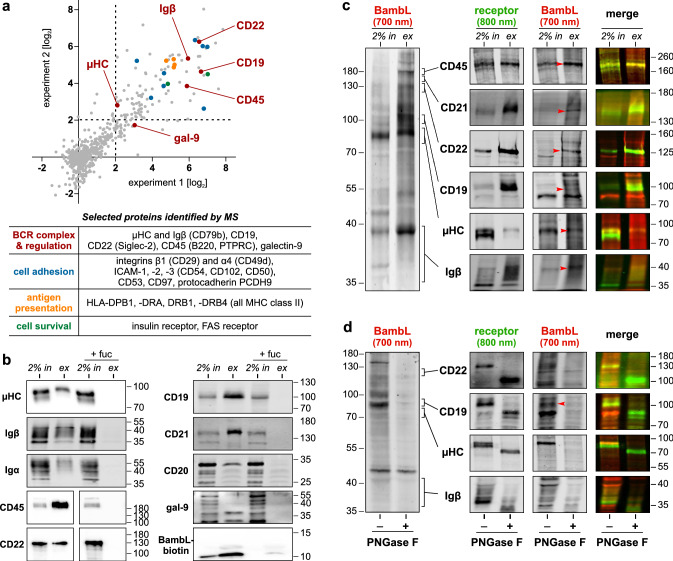
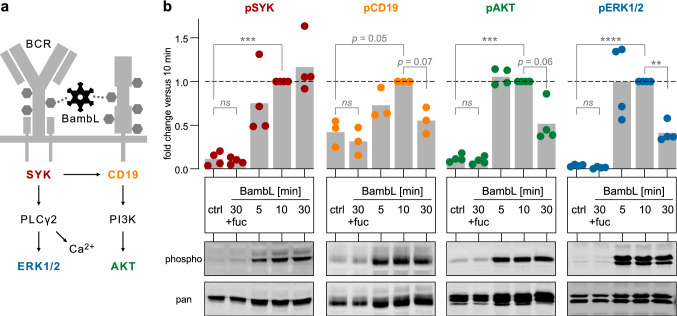
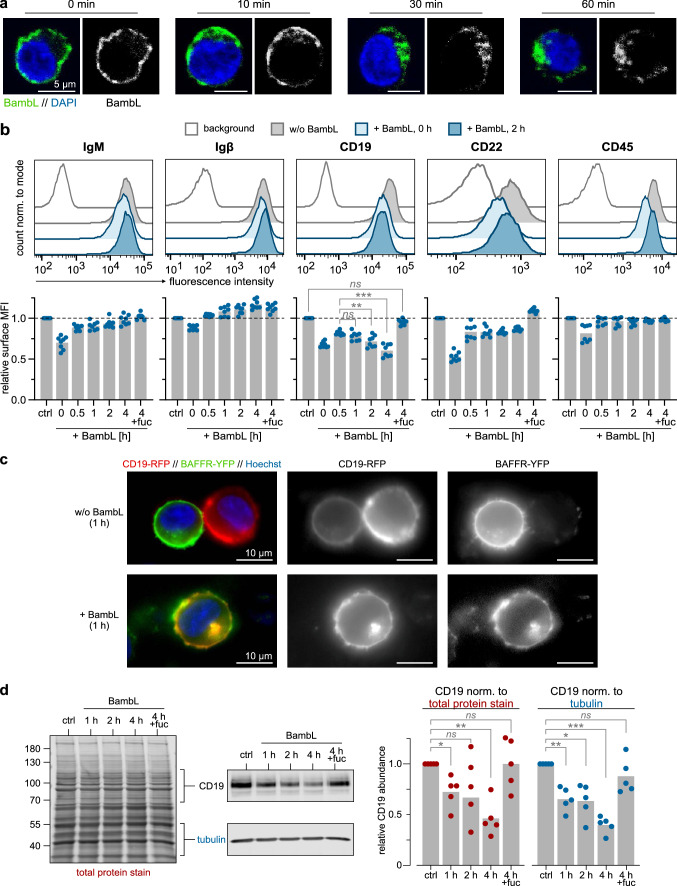
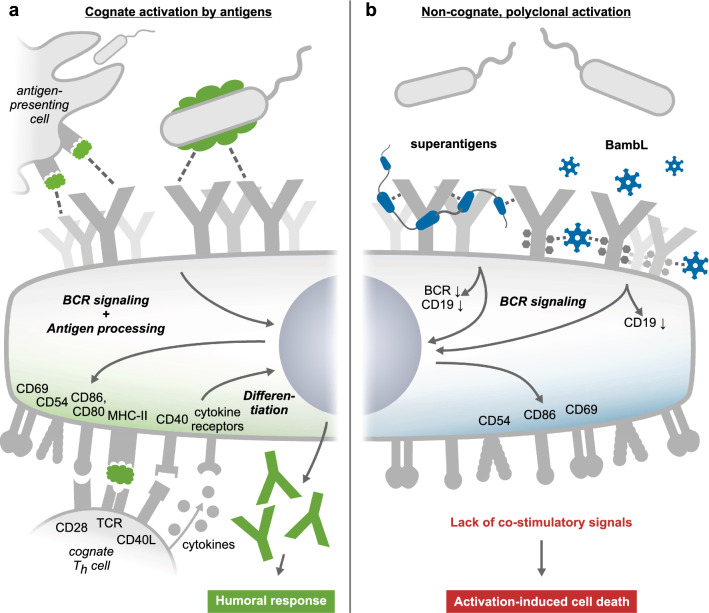
B cell superantigens crosslink conserved domains of B cell receptors (BCRs) and cause dysregulated, polyclonal B cell activation irrespective of normal BCR-antigen complementarity. The cells typically succumb to activation-induced cell death, which can impede the adaptive immune response and favor infection. In the present study, we demonstrate that the fucose-binding lectin of Burkholderia ambifaria, BambL, bears functional resemblance to B cell superantigens. By engaging surface glycans, the bacterial lectin activated human peripheral blood B cells, which manifested in the surface expression of CD69, CD54 and CD86 but became increasingly cytotoxic at higher concentrations. The effects were sensitive to BCR pathway inhibitors and excess fucose, which corroborates a glycan-driven mode of action. Interactome analyses in a model cell line suggest BambL binds directly to glycans of the BCR and regulatory coreceptors. In vitro, BambL triggered BCR signaling and induced CD19 internalization and degradation. Owing to the lectin's six binding sites, we propose a BCR activation model in which BambL functions as a clustering hub for receptor glycans, modulates normal BCR regulation, and induces cell death through exhaustive activation.
Keywords: Adaptive immunity; Apoptosis; Bacterial pathogens; Immunoglobulin glycosylation; Multivalent lectins; β-Propeller lectins.
© 2021. The Author(s).
Conflict of interest statement
The authors have no conflicts of interests to declare.
Figures
References
-
- Graille M, Stura EA, Housden NG, Beckingham JA, Bottomley SP, Beale D, Taussig MJ, Sutton BJ, Gore MG, Charbonnier J-B. Complex between Peptostreptococcus magnus protein L and a human antibody reveals structural convergence in the interaction modes of Fab binding proteins. Structure. 2001;9(8):679–687. doi: 10.1016/S0969-2126(01)00630-X. - DOI - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
