

Molecular and structural basis of olfactory sensory neuron axon coalescence by Kirrel receptors
- PMID: 34731636
- PMCID: PMC8628261
- DOI: 10.1016/j.celrep.2021.109940
Molecular and structural basis of olfactory sensory neuron axon coalescence by Kirrel receptors
Abstract
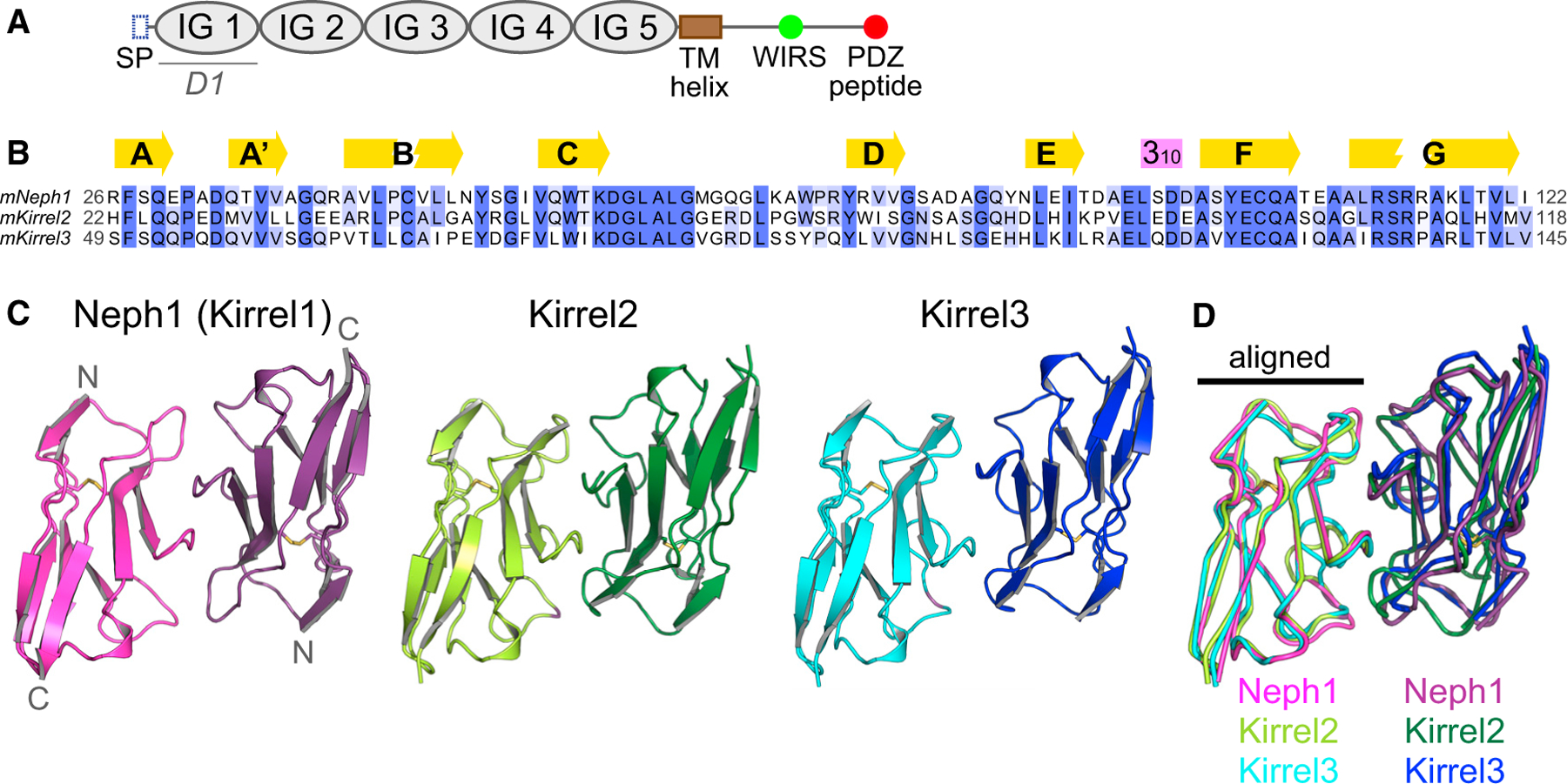
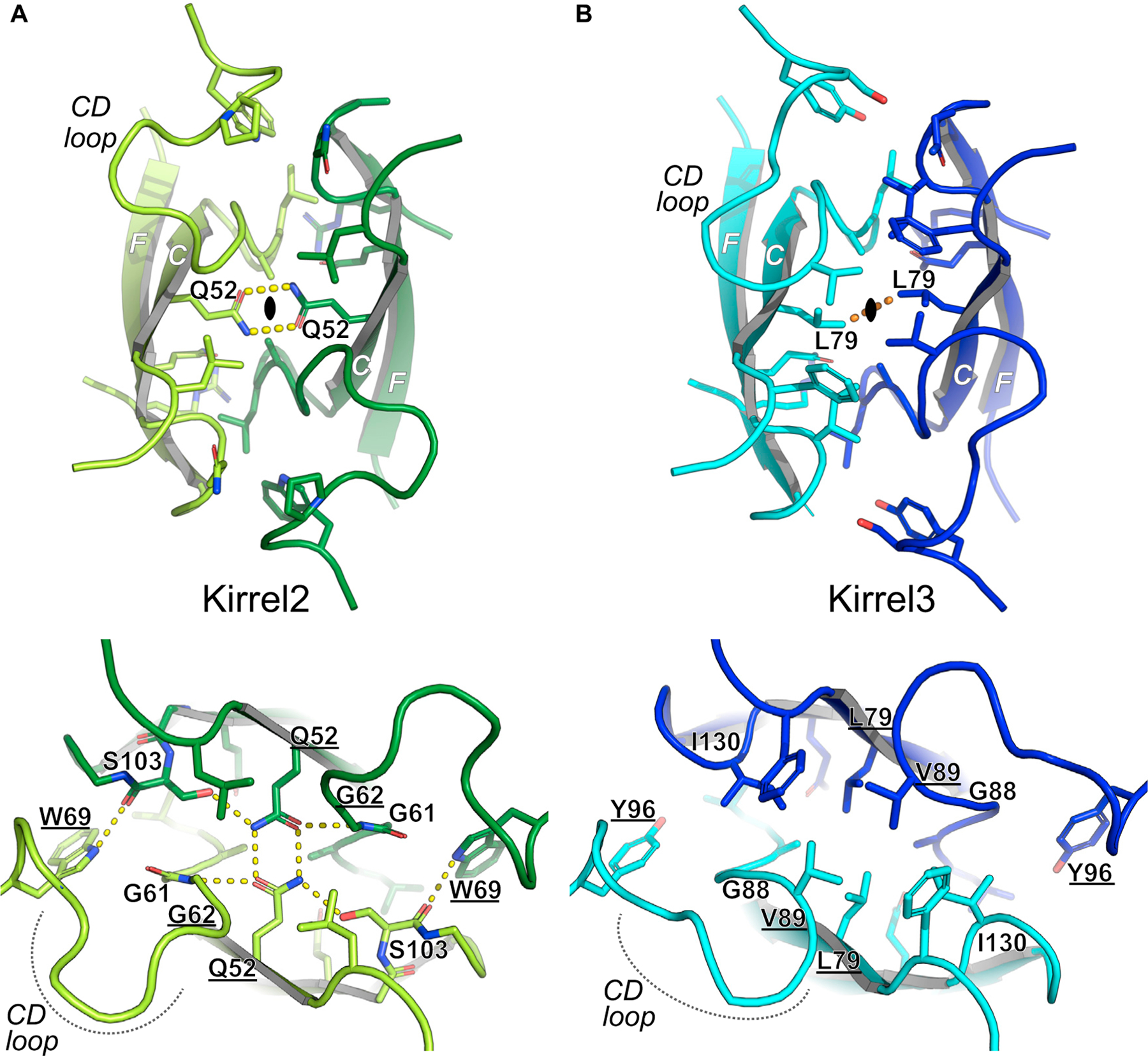
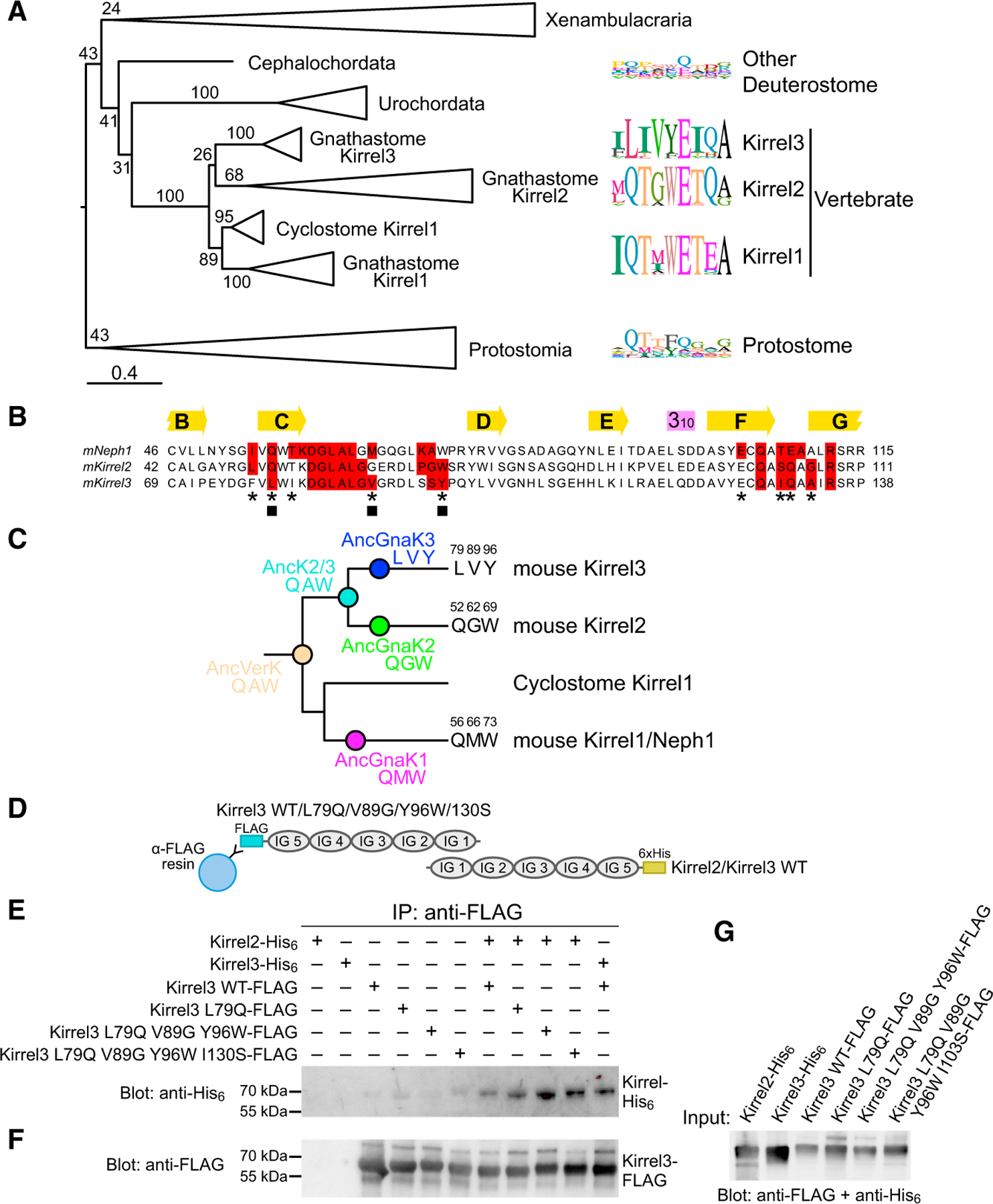
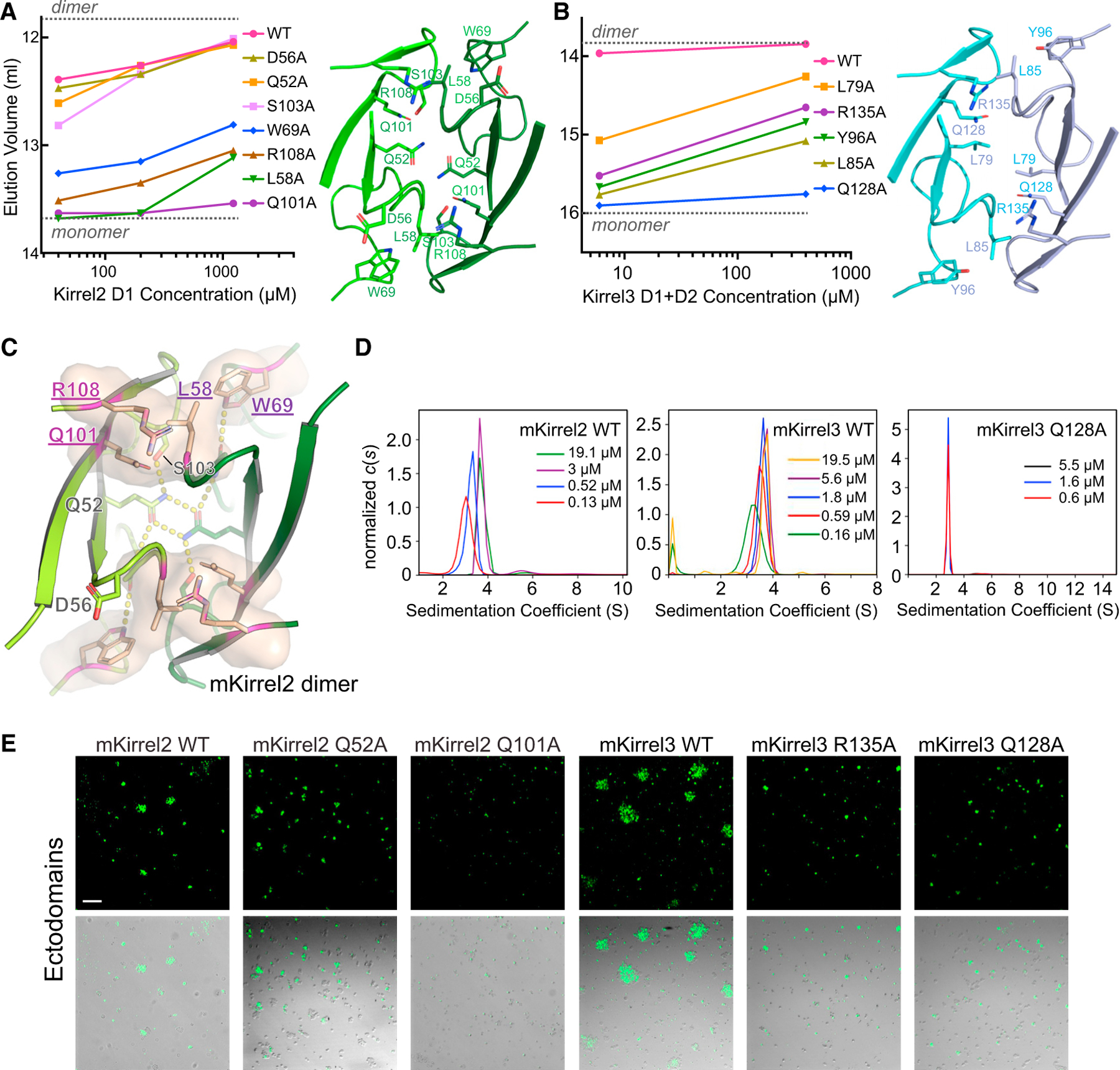
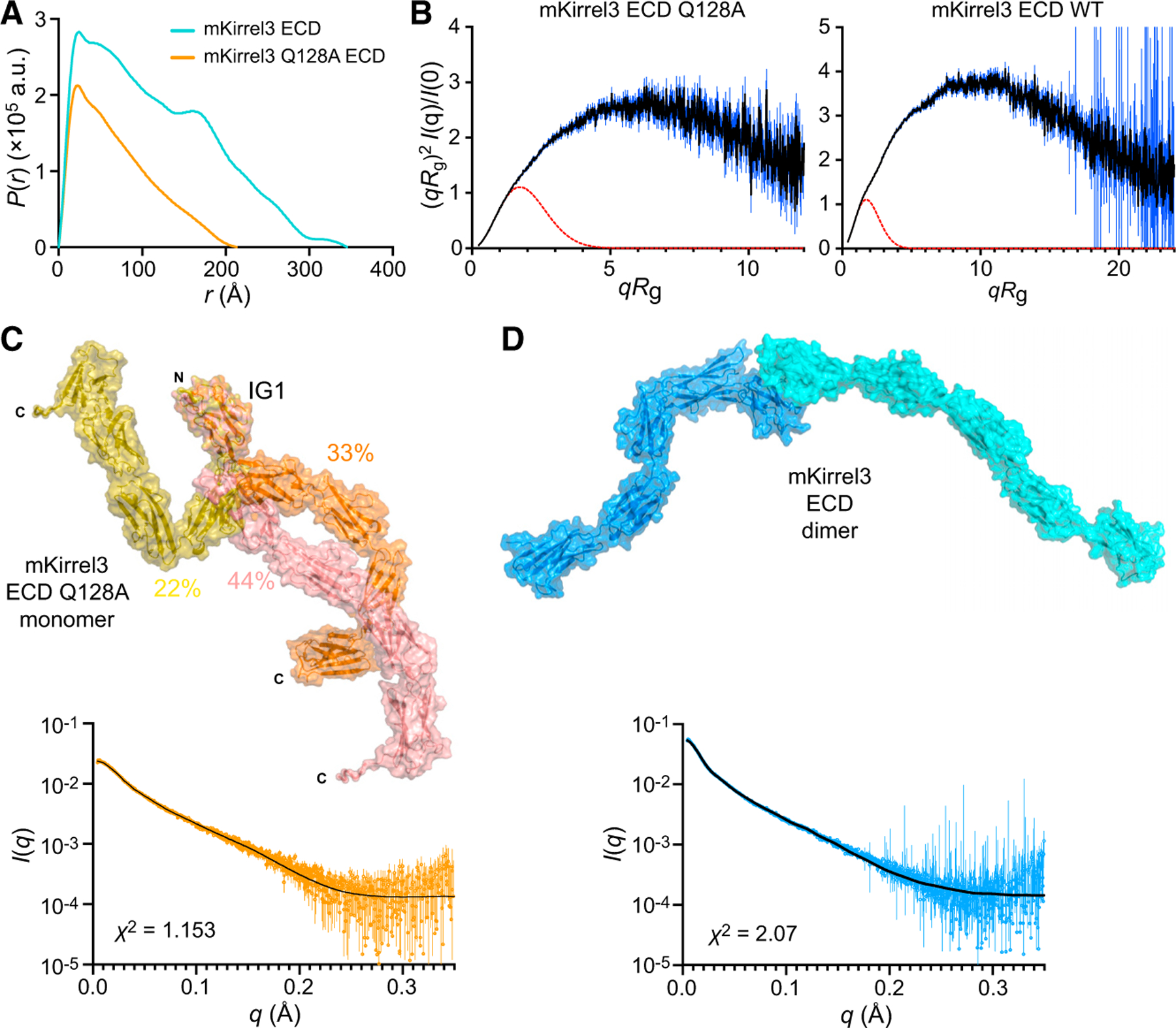
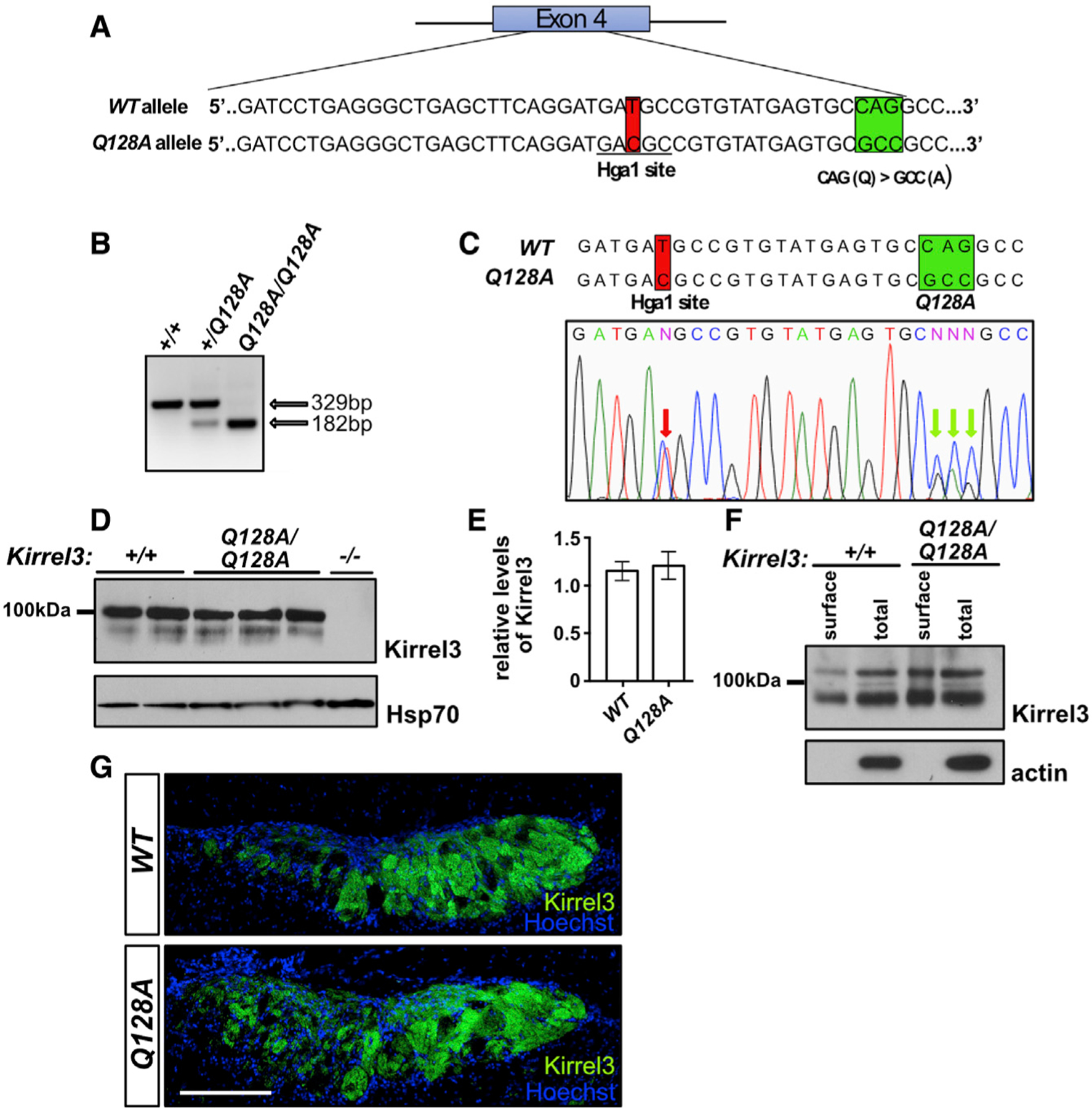
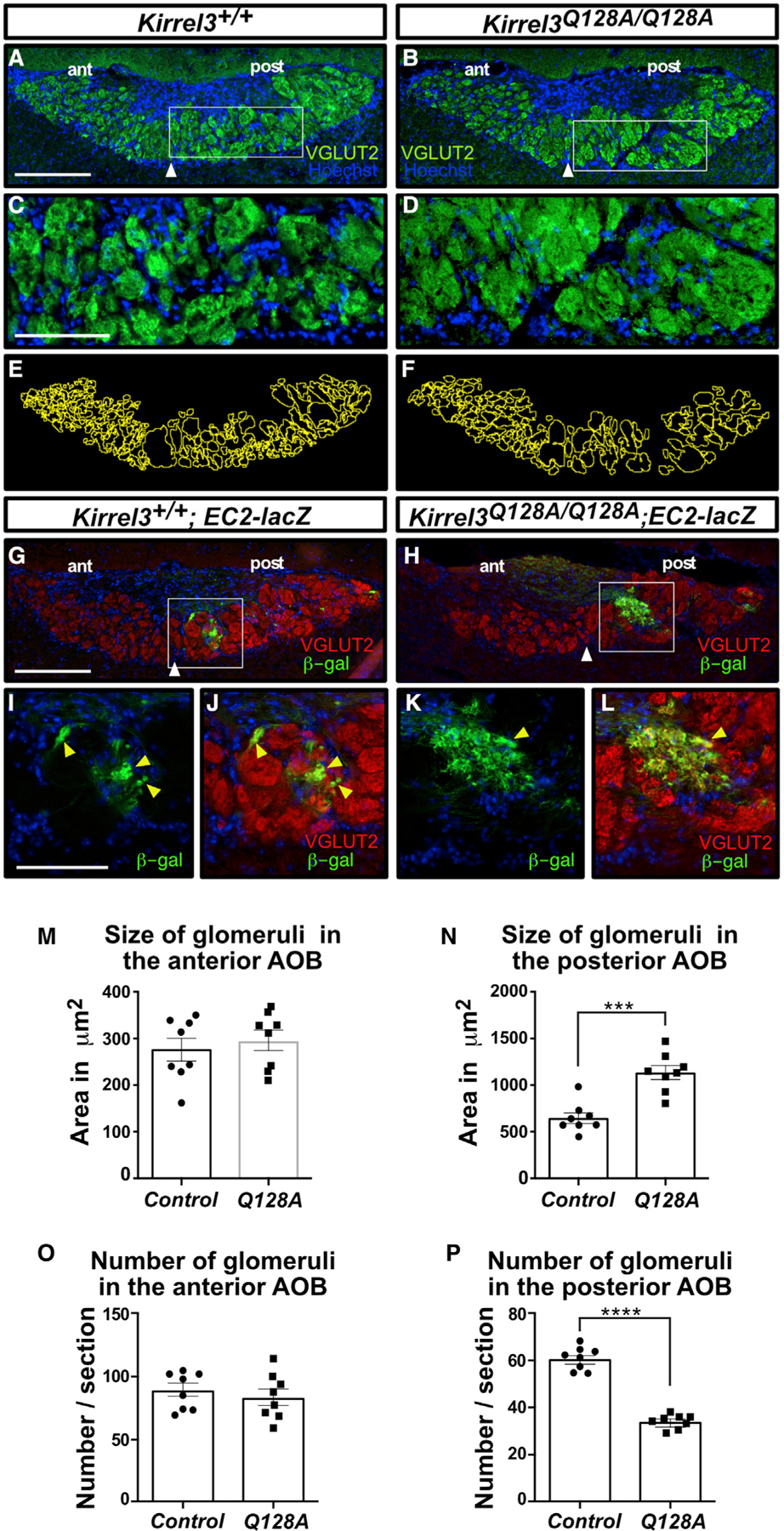
Projections from sensory neurons of olfactory systems coalesce into glomeruli in the brain. The Kirrel receptors are believed to homodimerize via their ectodomains and help separate sensory neuron axons into Kirrel2- or Kirrel3-expressing glomeruli. Here, we present the crystal structures of homodimeric Kirrel receptors and show that the closely related Kirrel2 and Kirrel3 have evolved specific sets of polar and hydrophobic interactions, respectively, disallowing heterodimerization while preserving homodimerization, likely resulting in proper segregation and coalescence of Kirrel-expressing axons into glomeruli. We show that the dimerization interface at the N-terminal immunoglobulin (IG) domains is necessary and sufficient to create homodimers and fail to find evidence for a secondary interaction site in Kirrel ectodomains. Furthermore, we show that abolishing dimerization of Kirrel3 in vivo leads to improper formation of glomeruli in the mouse accessory olfactory bulb as observed in Kirrel3-/- animals. Our results provide evidence for Kirrel3 homodimerization controlling axonal coalescence.
Keywords: Kirrel2; Kirrel3; accessory olfactory system; axonal coalescence; cell adhesion molecule; glomeruli; protein structure; vomeronasal.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
