

Striatal indirect pathway mediates exploration via collicular competition
- PMID: 34732888
- PMCID: PMC10281058
- DOI: 10.1038/s41586-021-04055-4
Striatal indirect pathway mediates exploration via collicular competition
Abstract
The ability to suppress actions that lead to a negative outcome and explore alternative actions is necessary for optimal decision making. Although the basal ganglia have been implicated in these processes1-5, the circuit mechanisms underlying action selection and exploration remain unclear. Here, using a simple lateralized licking task, we show that indirect striatal projection neurons (iSPN) in the basal ganglia contribute to these processes through modulation of the superior colliculus (SC). Optogenetic activation of iSPNs suppresses contraversive licking and promotes ipsiversive licking. Activity in lateral superior colliculus (lSC), a region downstream of the basal ganglia, is necessary for task performance and predicts lick direction. Furthermore, iSPN activation suppresses ipsilateral lSC, but surprisingly excites contralateral lSC, explaining the emergence of ipsiversive licking. Optogenetic inactivation reveals inter-collicular competition whereby each hemisphere of the superior colliculus inhibits the other, thus allowing the indirect pathway to disinhibit the contralateral lSC and trigger licking. Finally, inactivating iSPNs impairs suppression of devalued but previously rewarded licking and reduces exploratory licking. Our results reveal that iSPNs engage the competitive interaction between lSC hemispheres to trigger a motor action and suggest a general circuit mechanism for exploration during action selection.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Declaration of Interests
Dr. Sabatini is a founder and holds private equity in Optogenix. Tapered fibers that are commercially available from Optogenix were used as tools in the research.
Figures













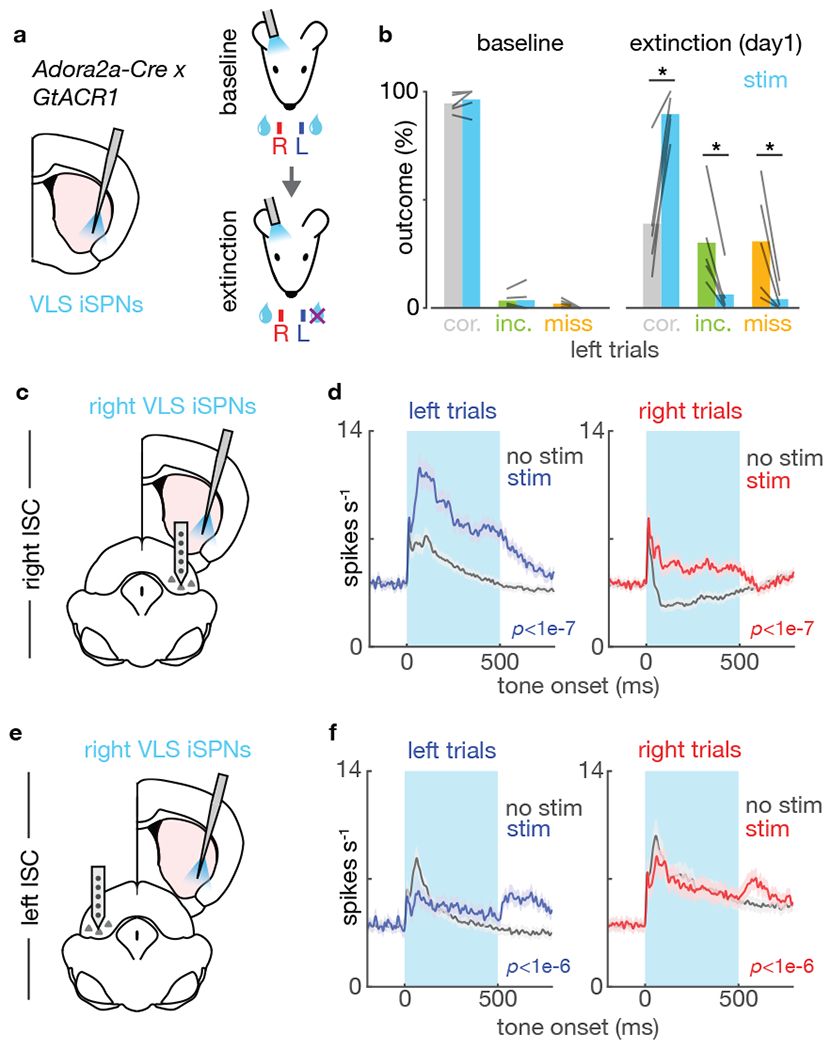
References
-
- Nonomura S et al. Monitoring and Updating of Action Selection for Goal-Directed Behavior through the Striatal Direct and Indirect Pathways. Neuron 99, 1302–1314.e5 (2018). - PubMed
Additional References
-
- Bro R, Acar E & Kolda TG Resolving the sign ambiguity in the singular value decomposition. J. Chemom 22, 135–140 (2008).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
