

The emerging role of ISWI chromatin remodeling complexes in cancer
- PMID: 34736517
- PMCID: PMC8567610
- DOI: 10.1186/s13046-021-02151-x
The emerging role of ISWI chromatin remodeling complexes in cancer
Abstract
Disordered chromatin remodeling regulation has emerged as an essential driving factor for cancers. Imitation switch (ISWI) family are evolutionarily conserved ATP-dependent chromatin remodeling complexes, which are essential for cellular survival and function through multiple genetic and epigenetic mechanisms. Omics sequencing and a growing number of basic and clinical studies found that ISWI family members displayed widespread gene expression and genetic status abnormalities in human cancer. Their aberrant expression is closely linked to patient outcome and drug response. Functional or componential alteration in ISWI-containing complexes is critical for tumor initiation and development. Furthermore, ISWI-non-coding RNA regulatory networks and some non-coding RNAs derived from exons of ISWI member genes play important roles in tumor progression. Therefore, unveiling the transcriptional regulation mechanism underlying ISWI family sparked a booming interest in finding ISWI-based therapies in cancer. This review aims at describing the current state-of-the-art in the role of ISWI subunits and complexes in tumorigenesis, tumor progression, immunity and drug response, and presenting deep insight into the physiological and pathological implications of the ISWI transcription machinery in cancers.
Keywords: Cofactors; ISWI family; Inhibitors; Transcription complexes; Tumor immunology.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
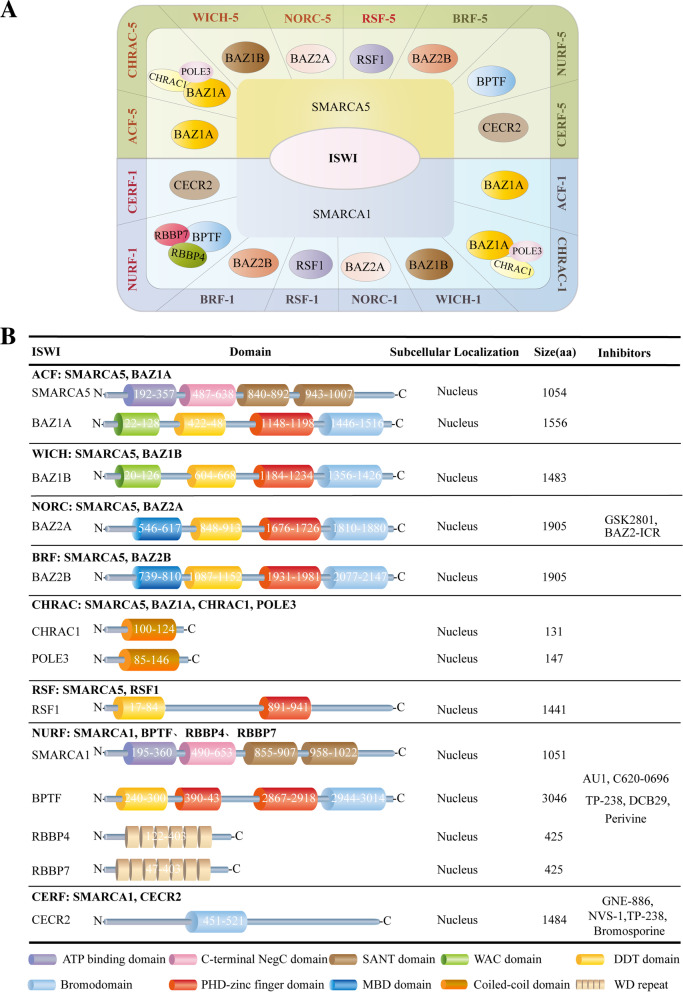
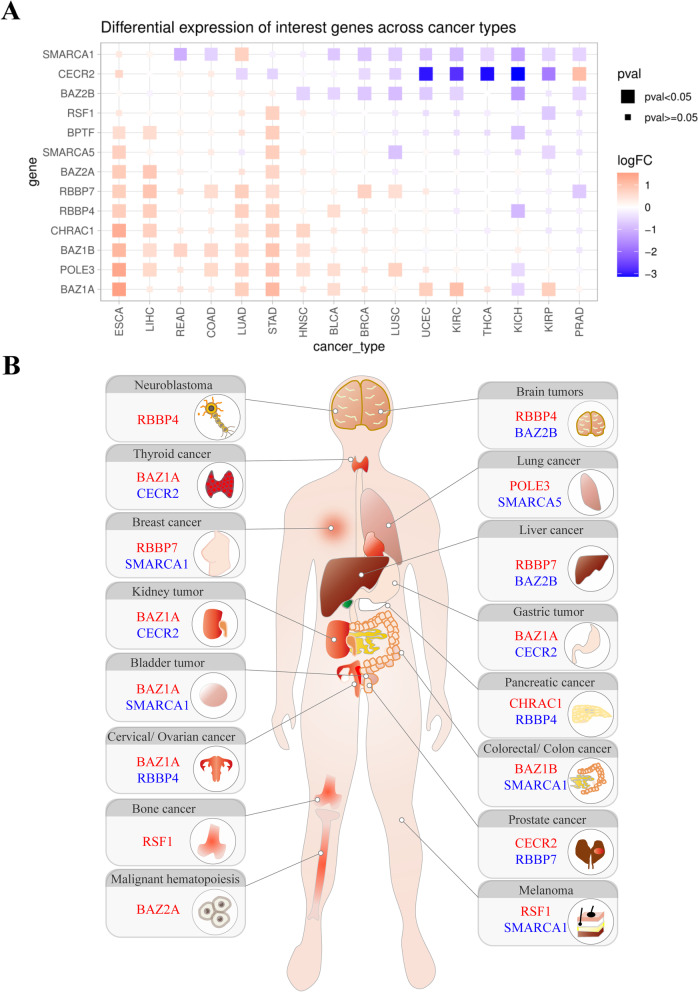
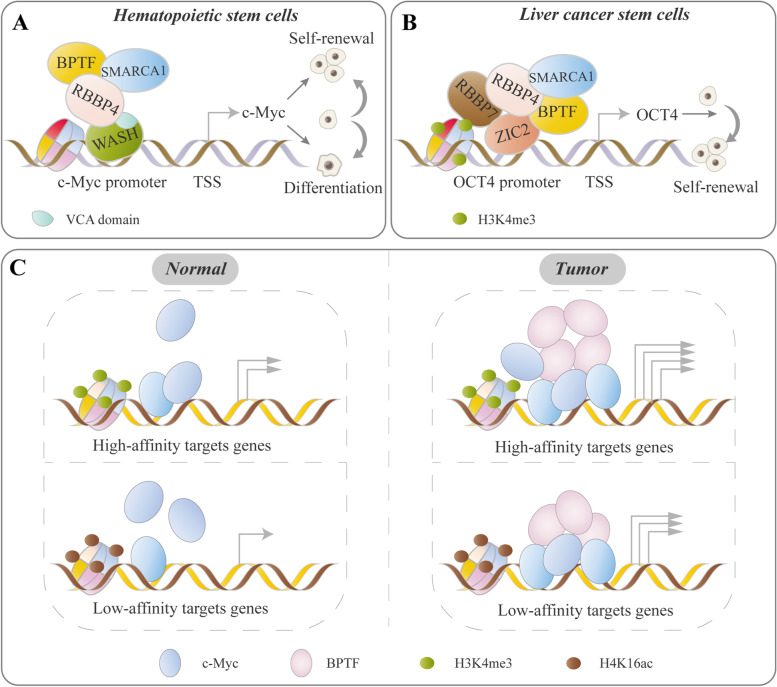
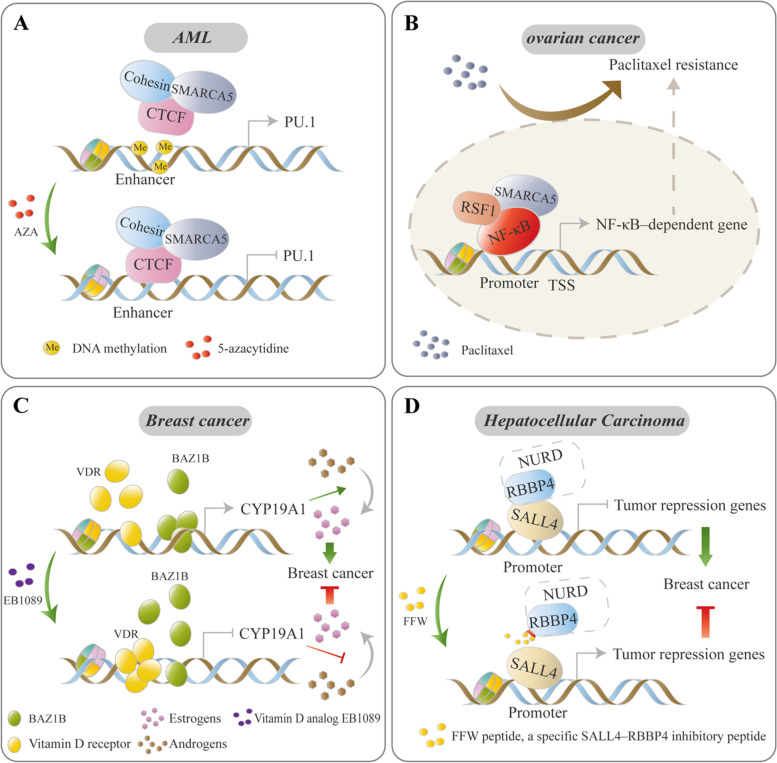
References
Publication types
MeSH terms
Substances
Grants and funding
- 2108YFA0107800/national key research and development program of china
- 81920108004, 81770107, 81470362/national natural science foundation of china
- 81702722/national natural science foundation of china
- BX201700292/national postdoctoral program for innovative talents
- 2018JJ3703/natural science foundation of hunan province
LinkOut - more resources
Full Text Sources
Medical
