

Ubiquitination is essential for recovery of cellular activities after heat shock
- PMID: 34739326
- PMCID: PMC8574219
- DOI: 10.1126/science.abc3593
Ubiquitination is essential for recovery of cellular activities after heat shock
Abstract
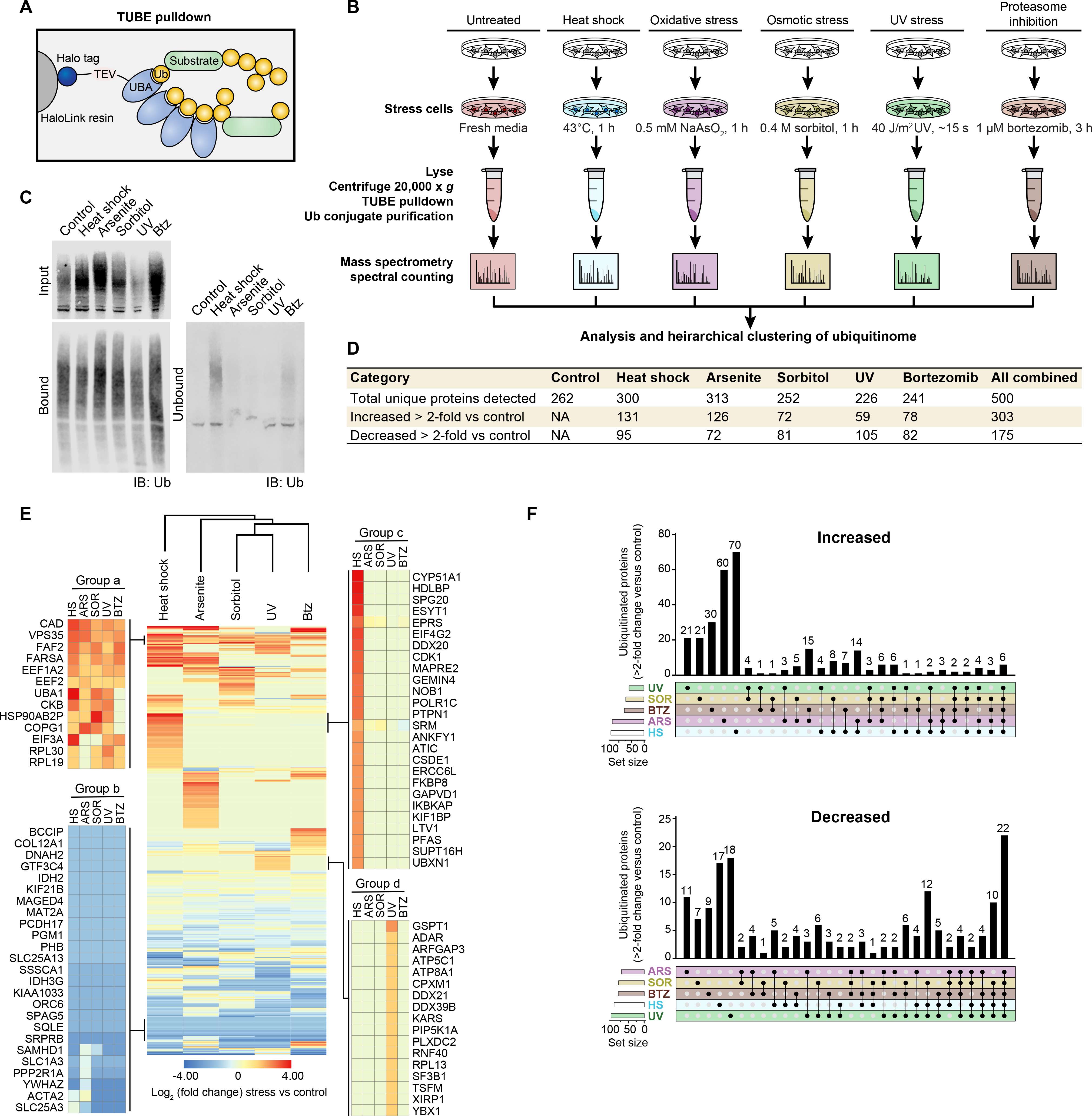
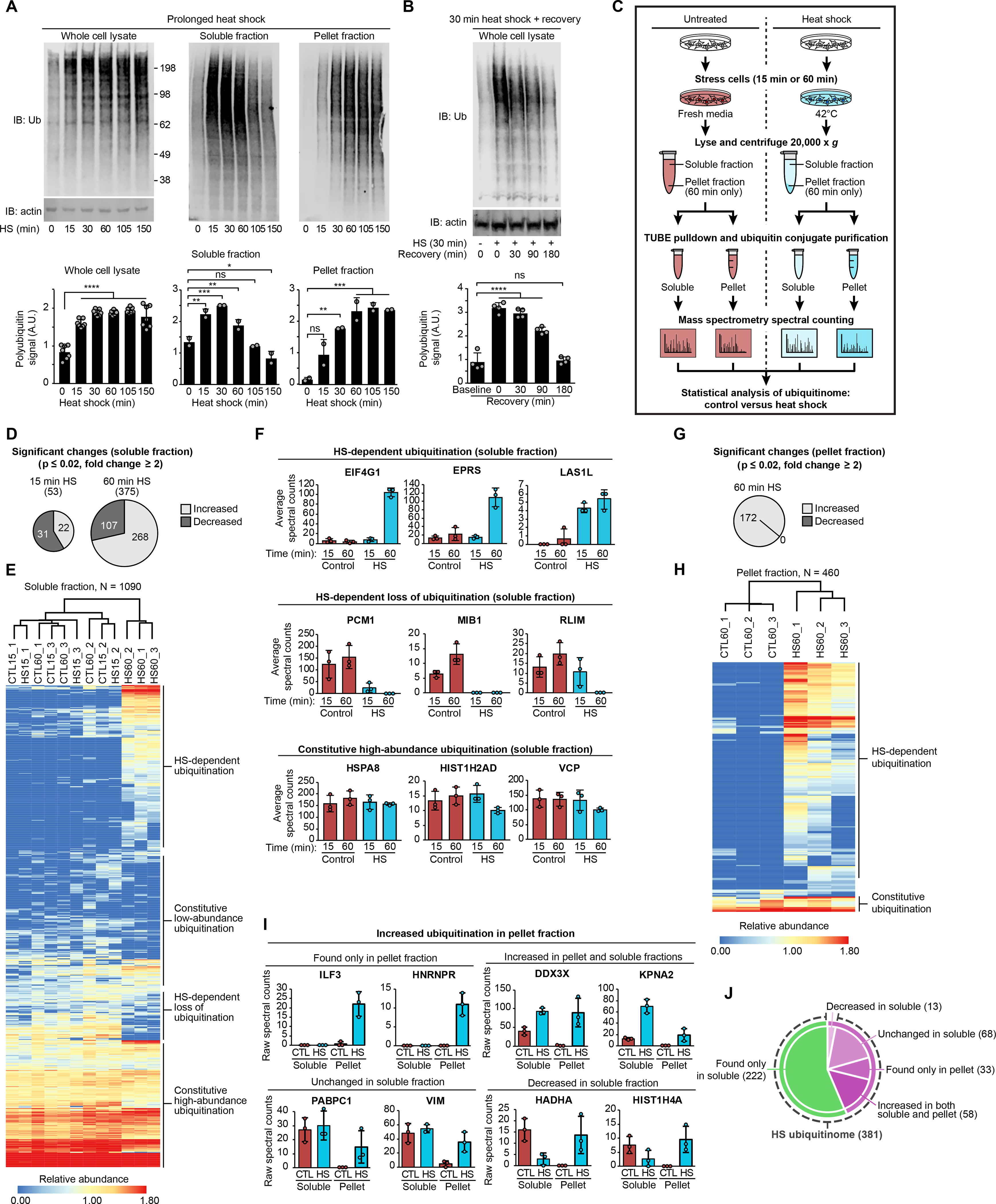
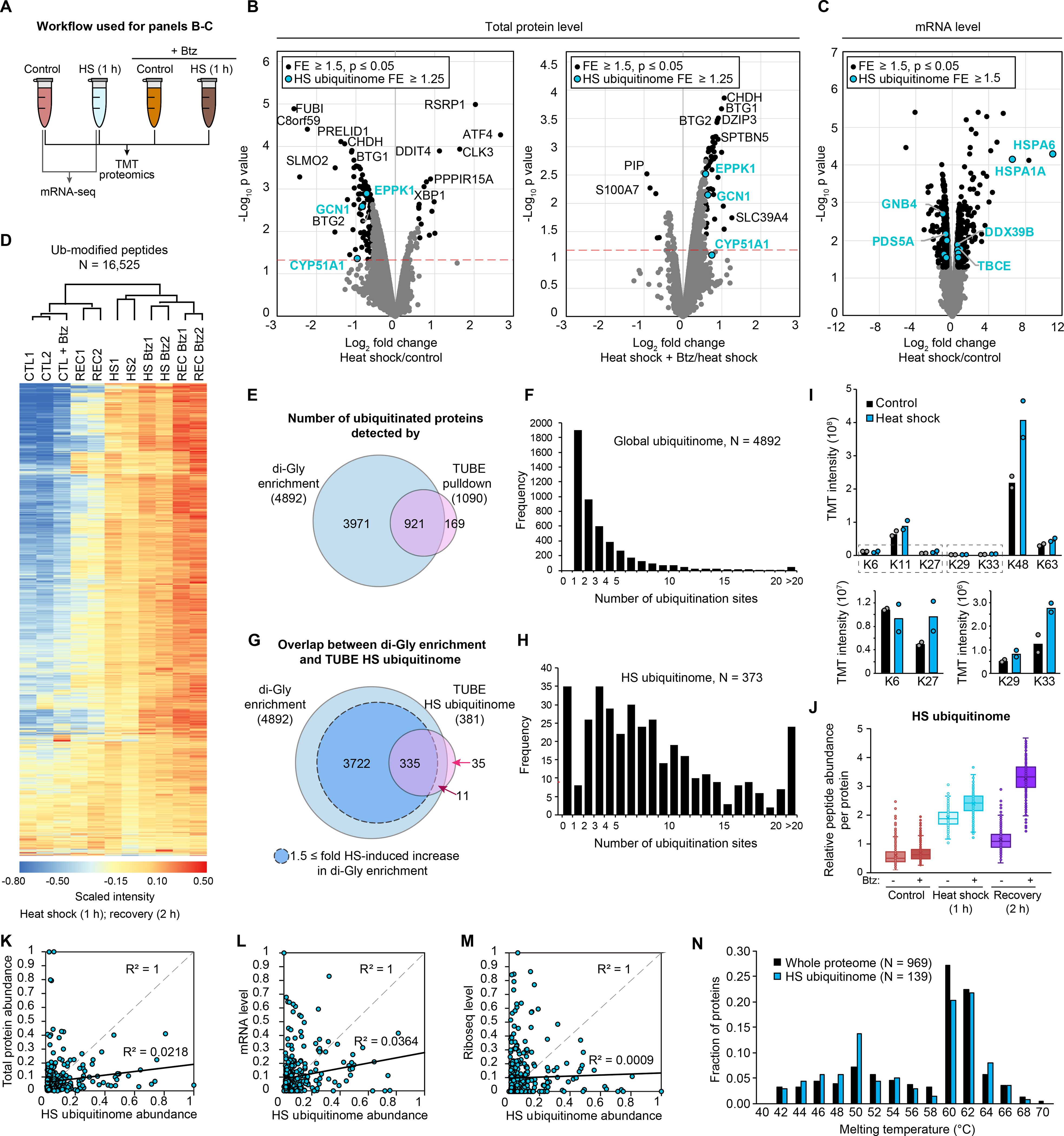
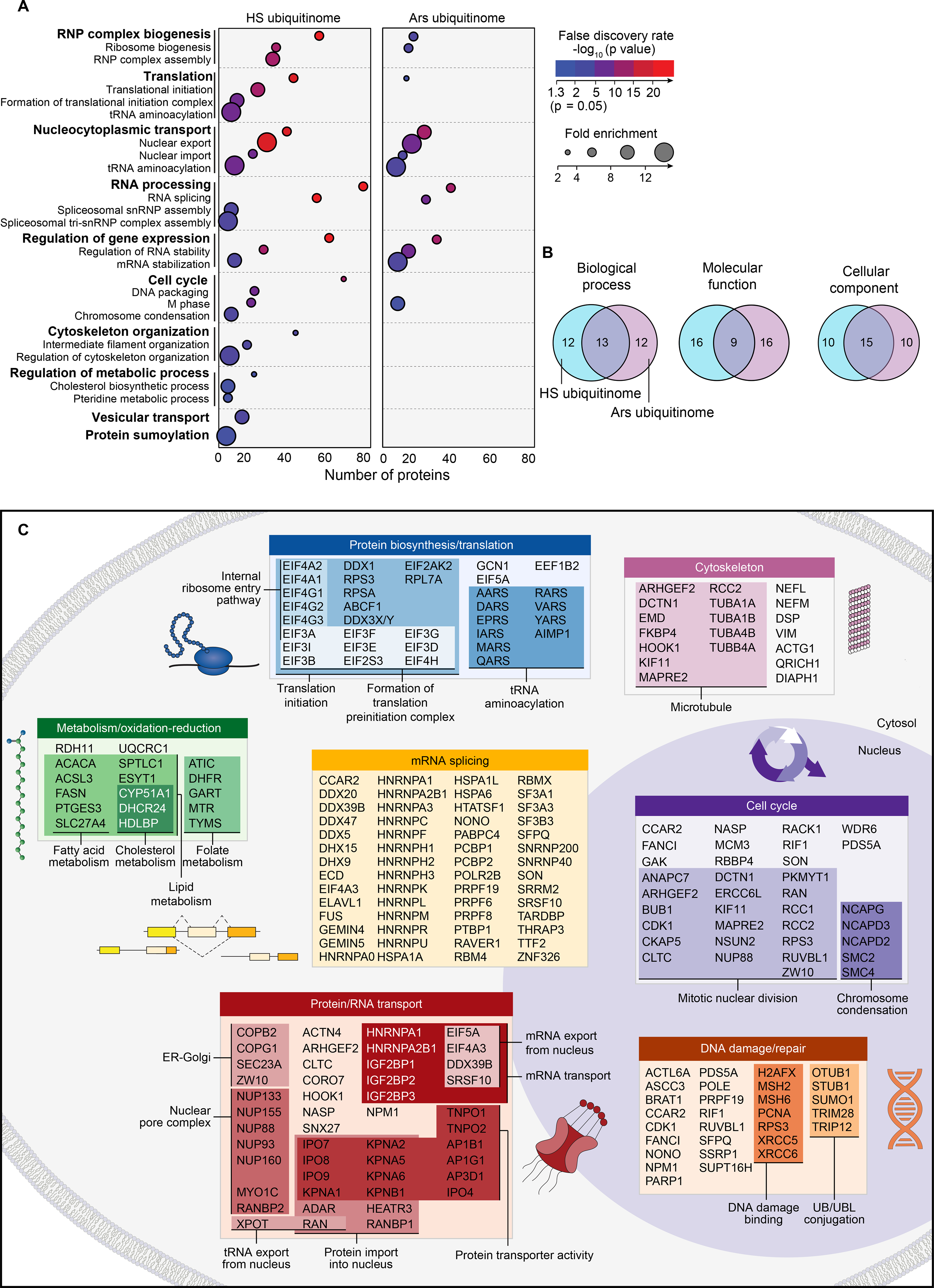
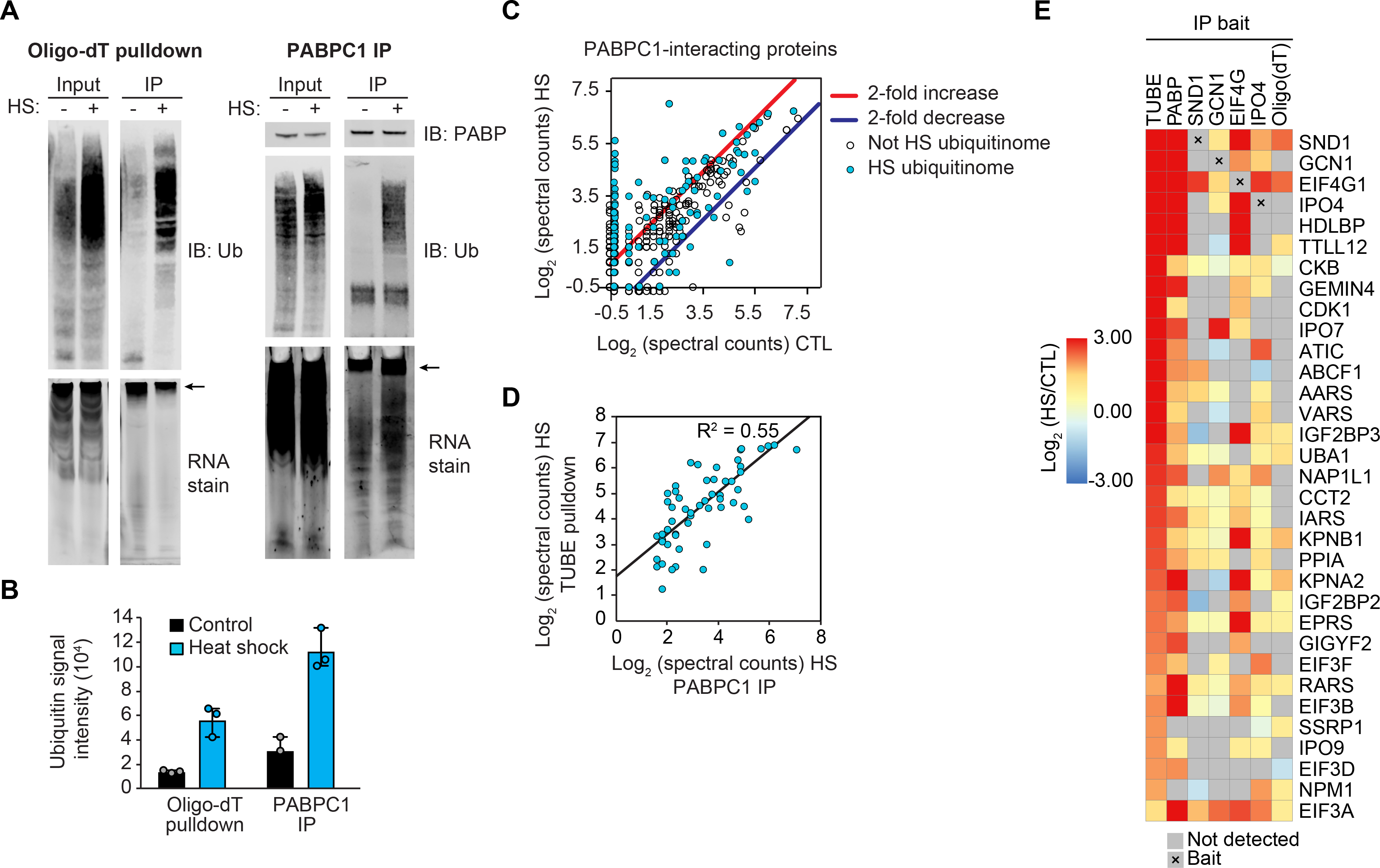
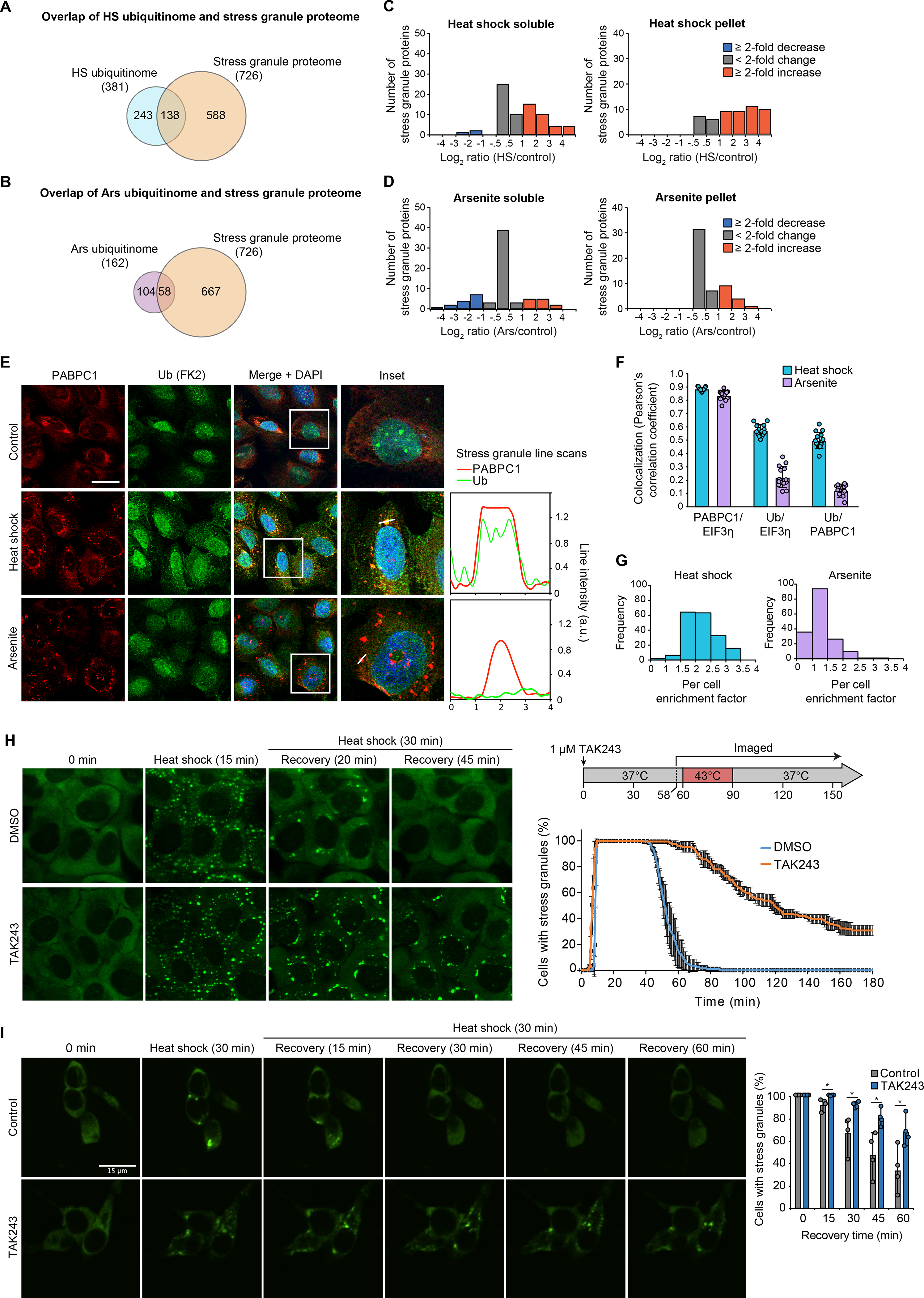
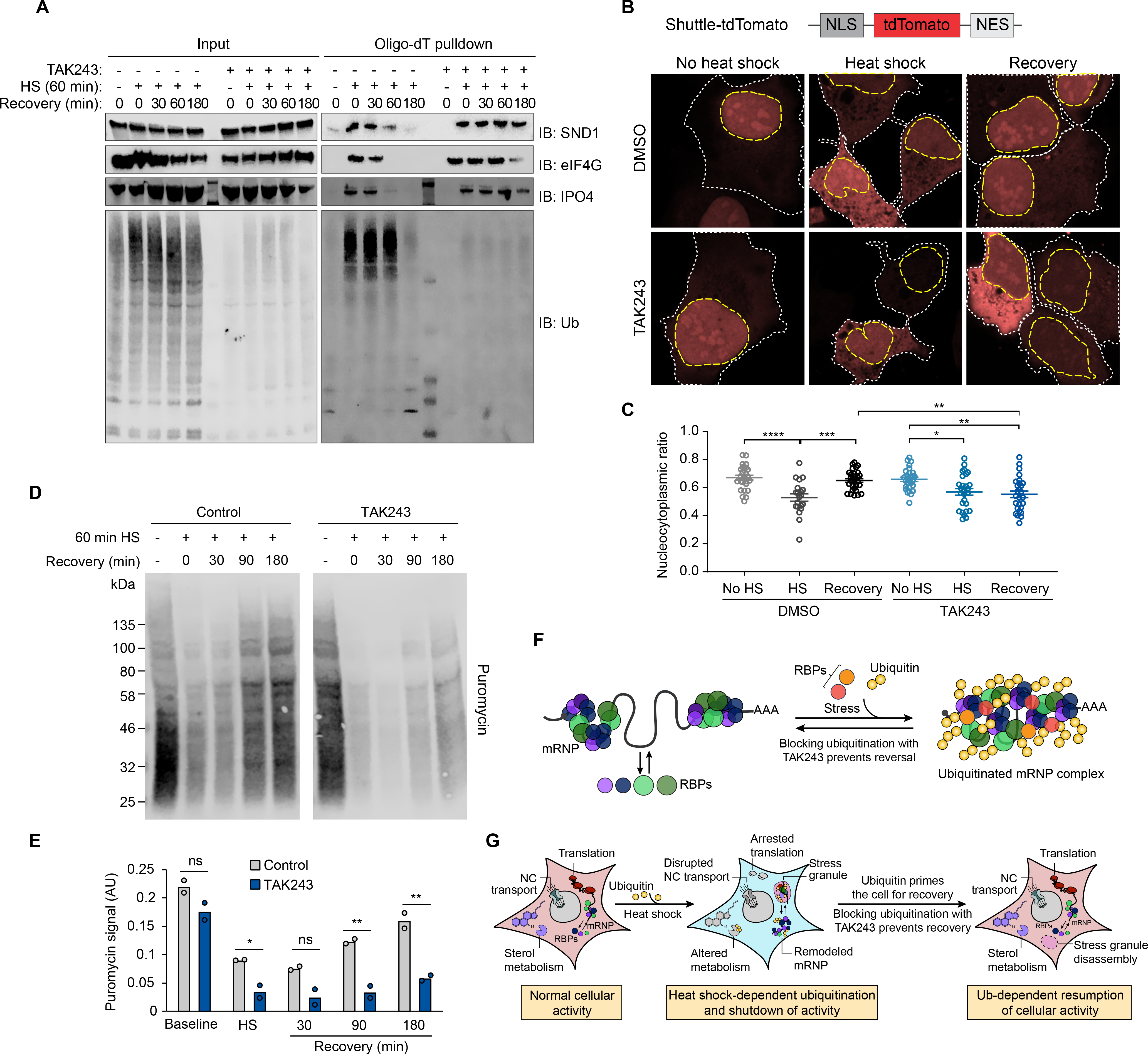
Eukaryotic cells respond to stress through adaptive programs that include reversible shutdown of key cellular processes, the formation of stress granules, and a global increase in ubiquitination. The primary function of this ubiquitination is thought to be for tagging damaged or misfolded proteins for degradation. Here, working in mammalian cultured cells, we found that different stresses elicited distinct ubiquitination patterns. For heat stress, ubiquitination targeted specific proteins associated with cellular activities that are down-regulated during stress, including nucleocytoplasmic transport and translation, as well as stress granule constituents. Ubiquitination was not required for the shutdown of these processes or for stress granule formation but was essential for the resumption of cellular activities and for stress granule disassembly. Thus, stress-induced ubiquitination primes the cell for recovery after heat stress.
Conflict of interest statement
Figures
Comment in
-
Managing stress granule disassembly with ubiquitin and its cousin.Signal Transduct Target Ther. 2021 Nov 11;6(1):391. doi: 10.1038/s41392-021-00782-2. Signal Transduct Target Ther. 2021. PMID: 34764249 Free PMC article. No abstract available.
References
-
- Harding HP et al. , An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell 11, 619–633 (2003). - PubMed
-
- Brostrom CO, Prostko CR, Kaufman RJ, Brostrom MA, Inhibition of translational initiation by activators of the glucose-regulated stress protein and heat shock protein stress response systems. Role of the interferon-inducible double-stranded RNA-activated eukaryotic initiation factor 2alpha kinase. J Biol Chem 271, 24995–25002 (1996). - PubMed
-
- Dever TE et al. , Phosphorylation of initiation factor 2 alpha by protein kinase GCN2 mediates gene-specific translational control of GCN4 in yeast. Cell 68, 585–596 (1992). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
