

A synthetic gene circuit for imaging-free detection of signaling pulses
- PMID: 34739875
- PMCID: PMC8857027
- DOI: 10.1016/j.cels.2021.10.002
A synthetic gene circuit for imaging-free detection of signaling pulses
Abstract
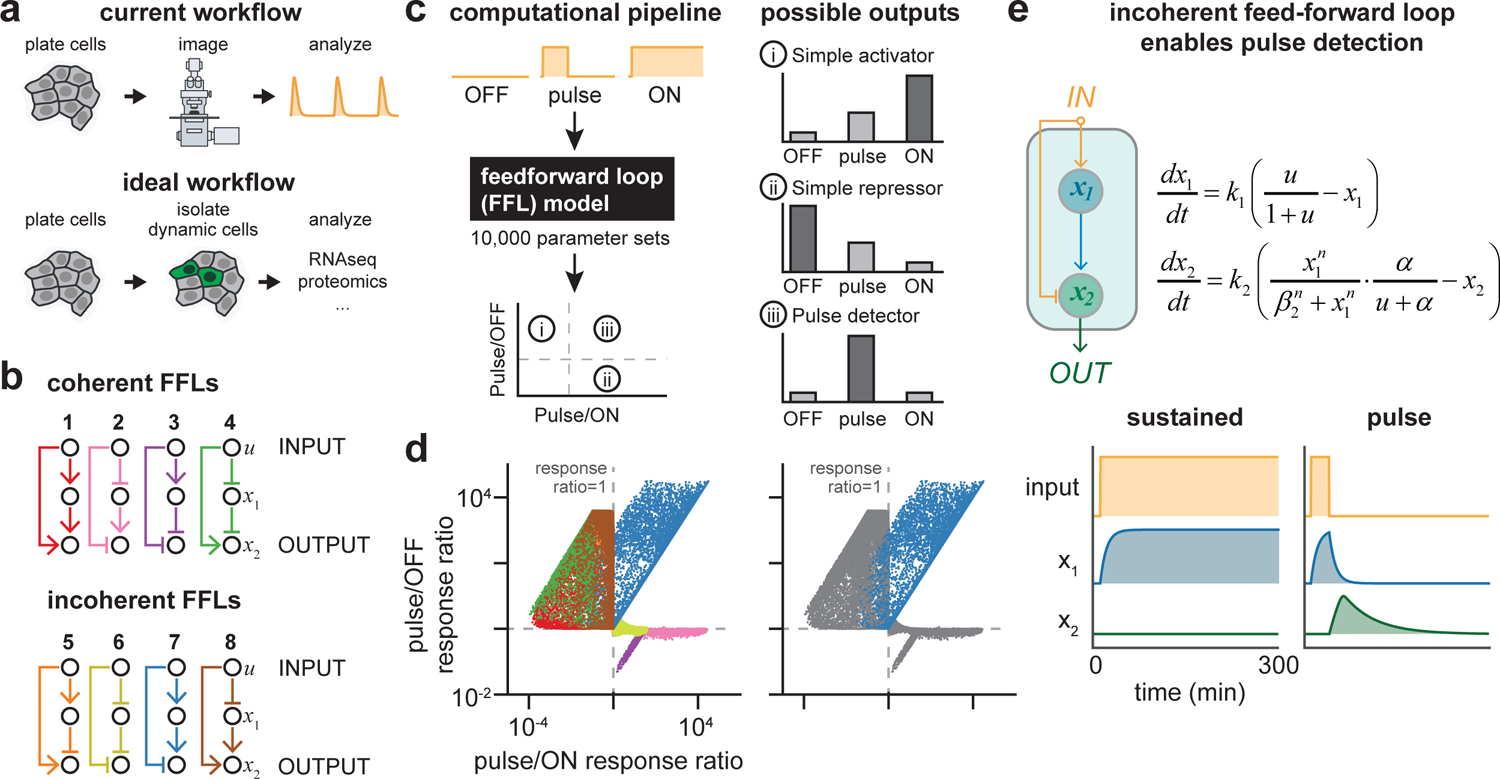
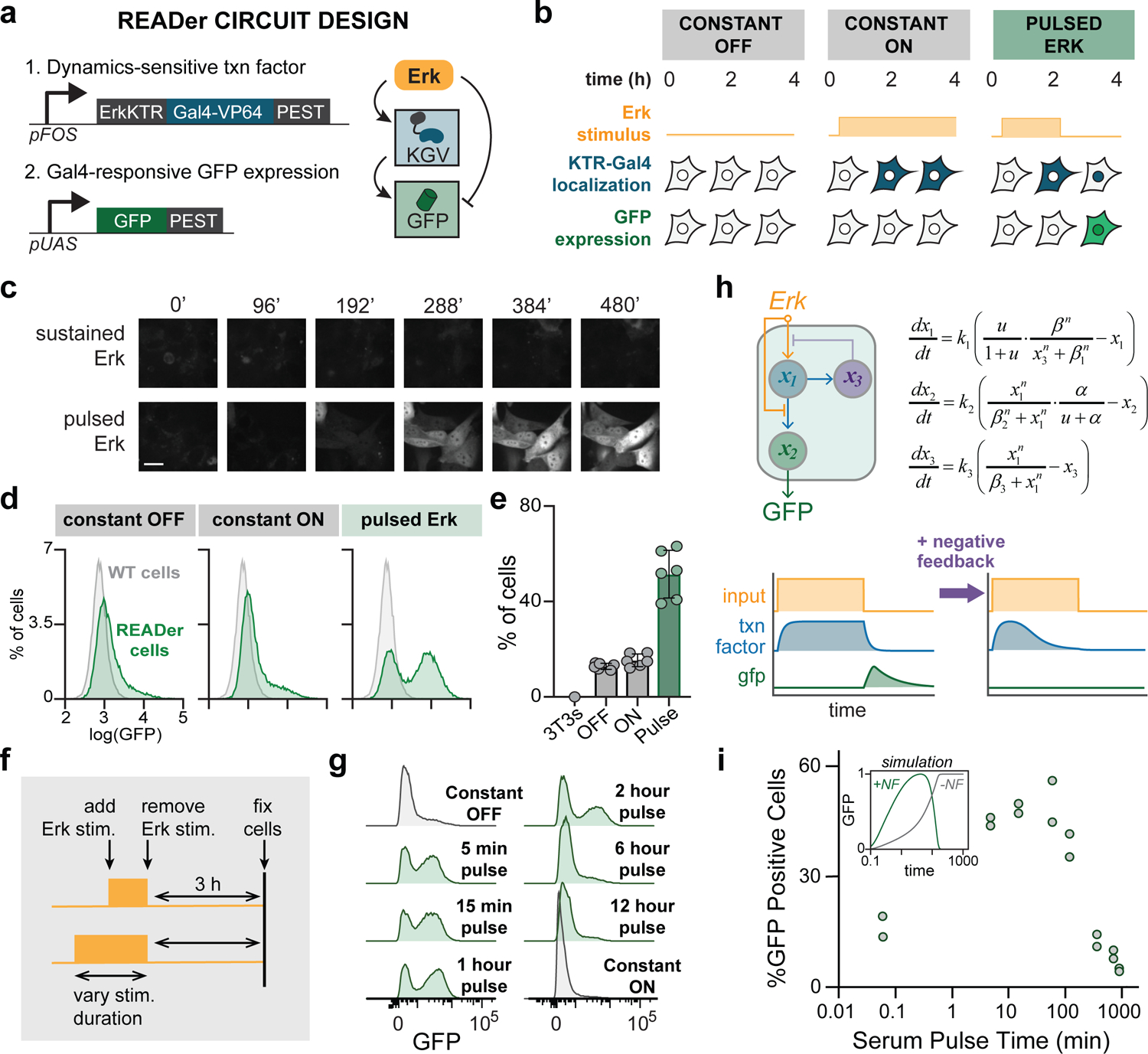
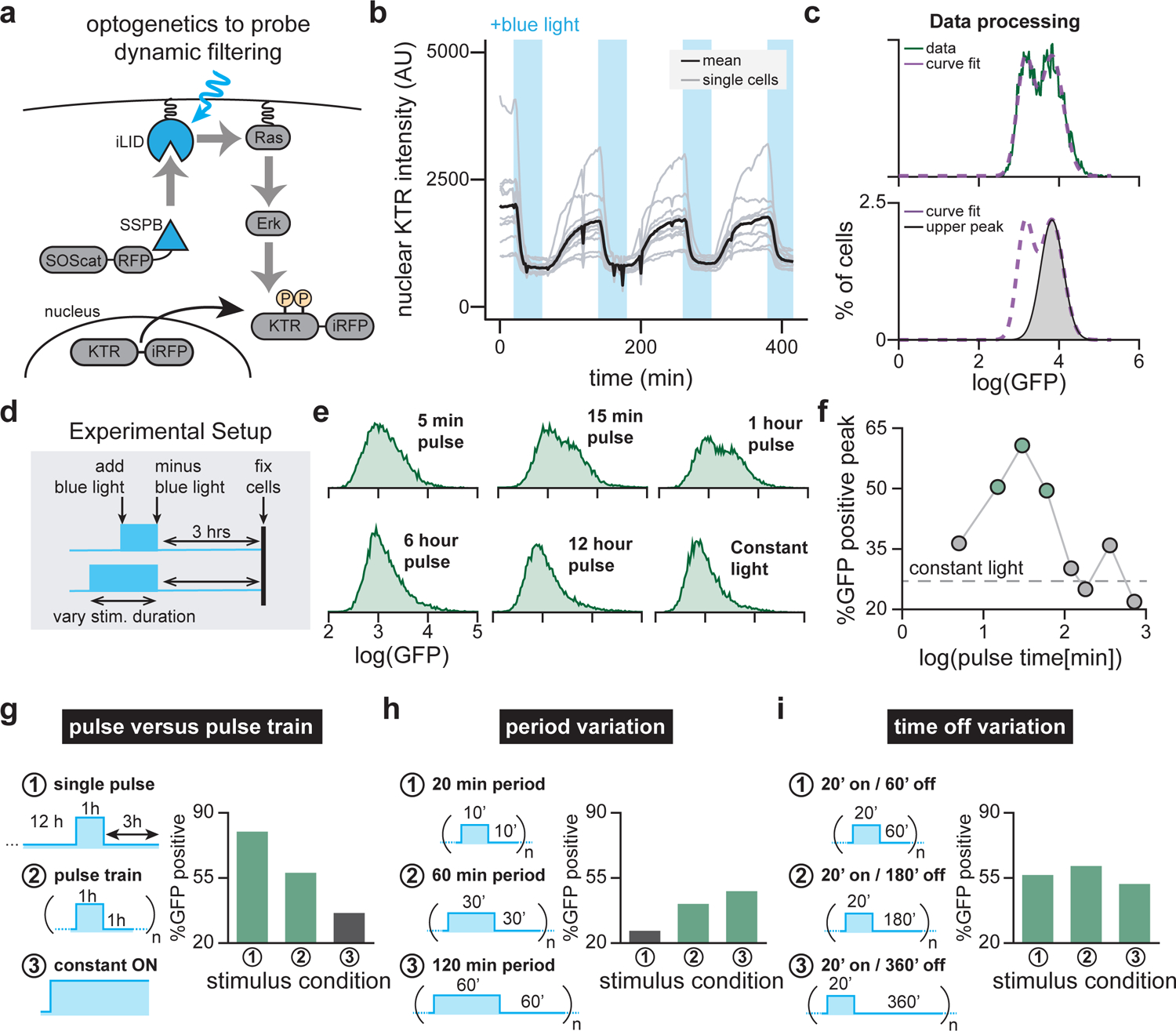
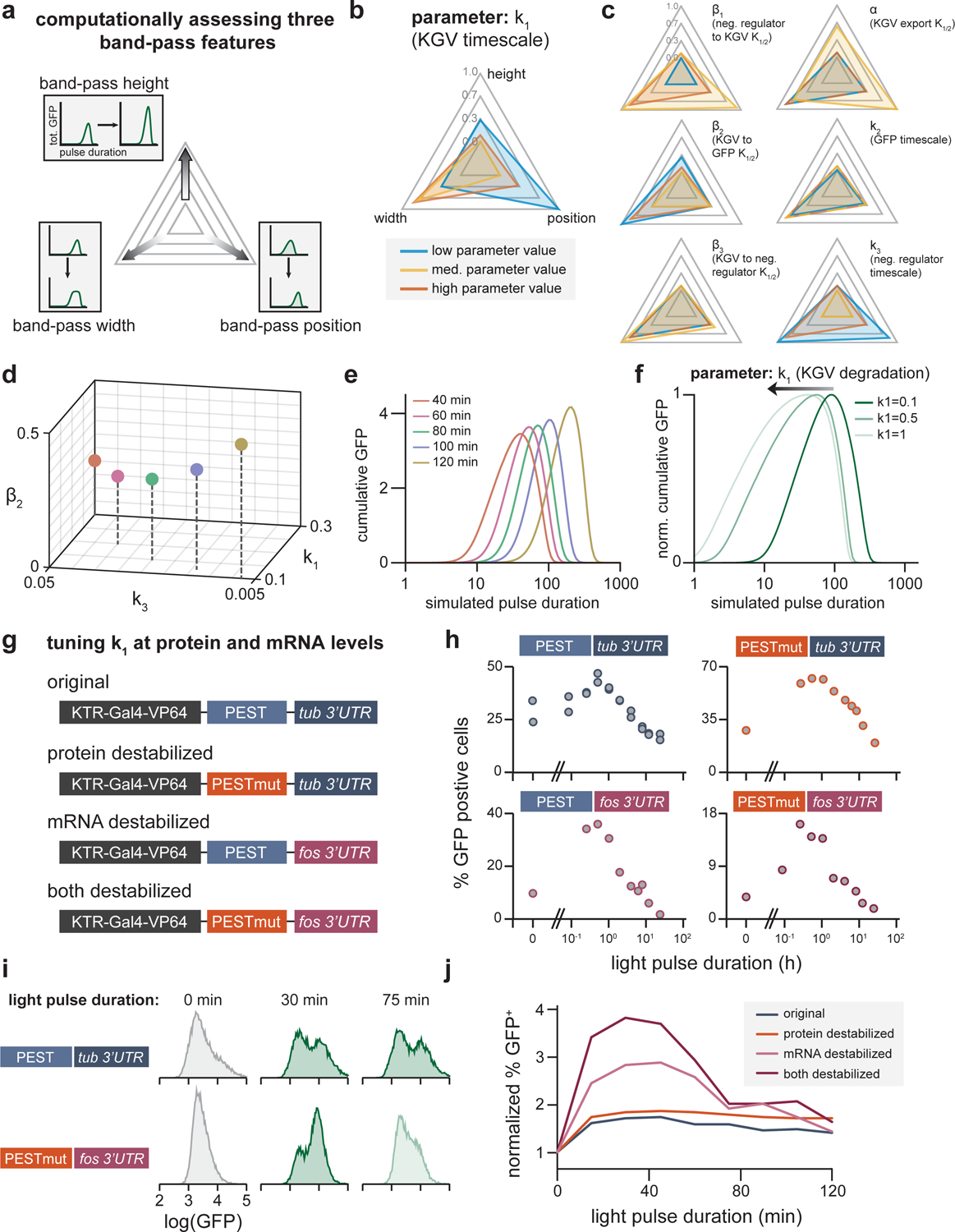
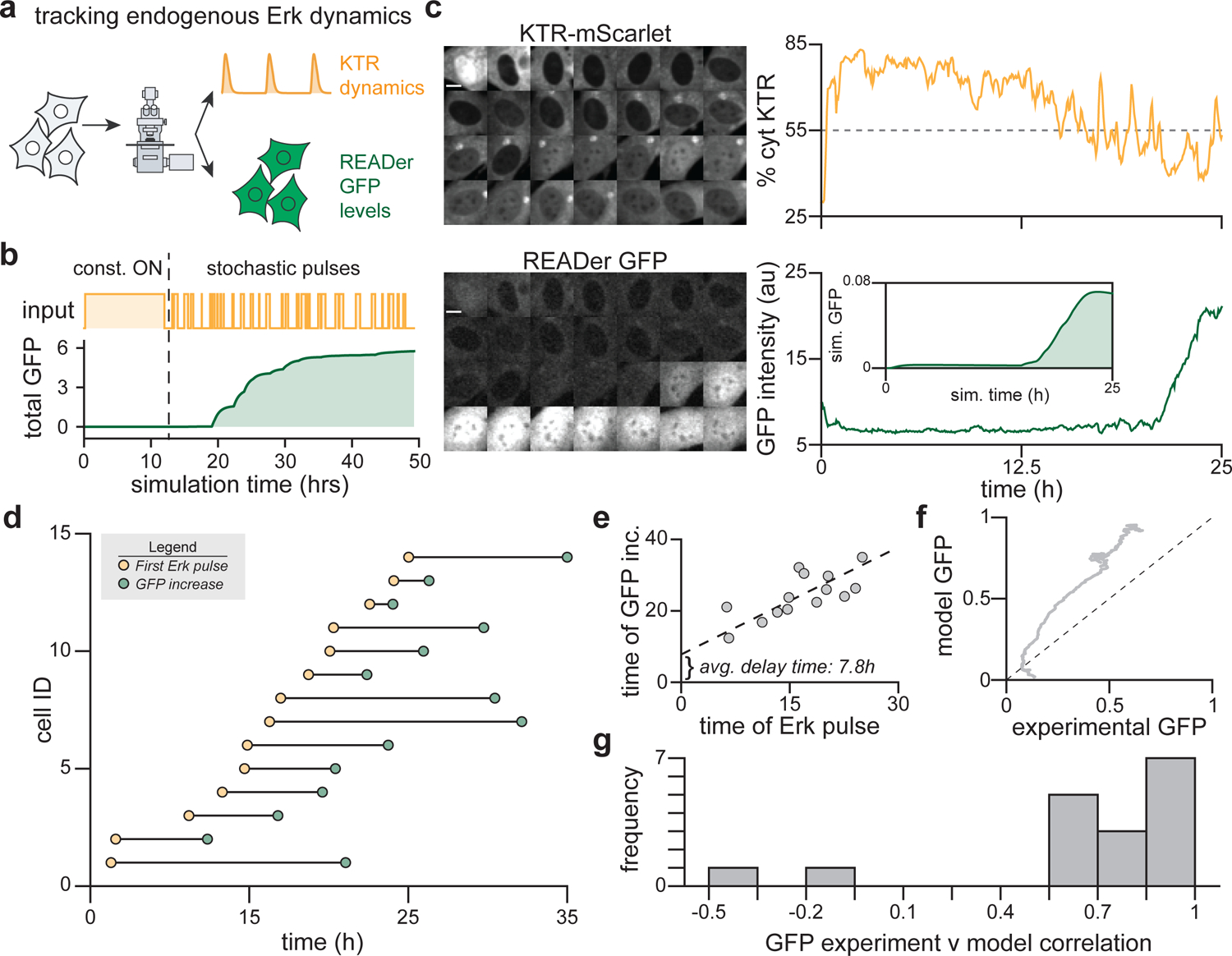
Cells employ intracellular signaling pathways to sense and respond to changes in their external environment. In recent years, live-cell biosensors have revealed complex pulsatile dynamics in many pathways, but studies of these signaling dynamics are limited by the necessity of live-cell imaging at high spatiotemporal resolution. Here, we describe an approach to infer pulsatile signaling dynamics from a single measurement in fixed cells using a pulse-detecting gene circuit. We computationally screened for circuits with the capability to selectively detect signaling pulses, revealing an incoherent feedforward topology that robustly performs this computation. We implemented the motif experimentally for the Erk signaling pathway using a single engineered transcription factor and fluorescent protein reporter. Our "recorder of Erk activity dynamics" (READer) responds sensitively to spontaneous and stimulus-driven Erk pulses. READer circuits open the door to permanently labeling transient, dynamic cell populations to elucidate the mechanistic underpinnings and biological consequences of signaling dynamics.
Keywords: cell signaling; network motifs; signaling dynamics; synthetic biology.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests J.E.T. is a member of the advisory board of Cell Systems. The authors declare no competing interests.
Figures

Comment in
-
Evaluation of Ravindran et al.: Real-time detection of signaling pulses in vivo: making cells monitor themselves.Cell Syst. 2022 Feb 16;13(2):105-106. doi: 10.1016/j.cels.2022.01.001. Cell Syst. 2022. PMID: 35176231
References
-
- Adler M, Szekely P, Mayo A & Alon U Optimal Regulatory Circuit Topologies for Fold-Change Detection. Cell Syst. 4, 171–181.e8 (2017). - PubMed
-
- Alon U (2007). Network motifs: theory and experimental approaches. Nat Rev Genet 8, 450–461. - PubMed
-
- Aoki K, Kumagai Y, Sakurai A, Komatsu N, Fujita Y, Shionyu C, and Matsuda M (2013). Stochastic ERK activation induced by noise and cell-to-cell propagation regulates cell density-dependent proliferation. Mol Cell 52, 529–540. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources