

Taxonomically and Functionally Distinct Ciliophora Assemblages Inhabiting Baltic Sea Ice
- PMID: 34748071
- PMCID: PMC9747827
- DOI: 10.1007/s00248-021-01915-4
Taxonomically and Functionally Distinct Ciliophora Assemblages Inhabiting Baltic Sea Ice
Abstract
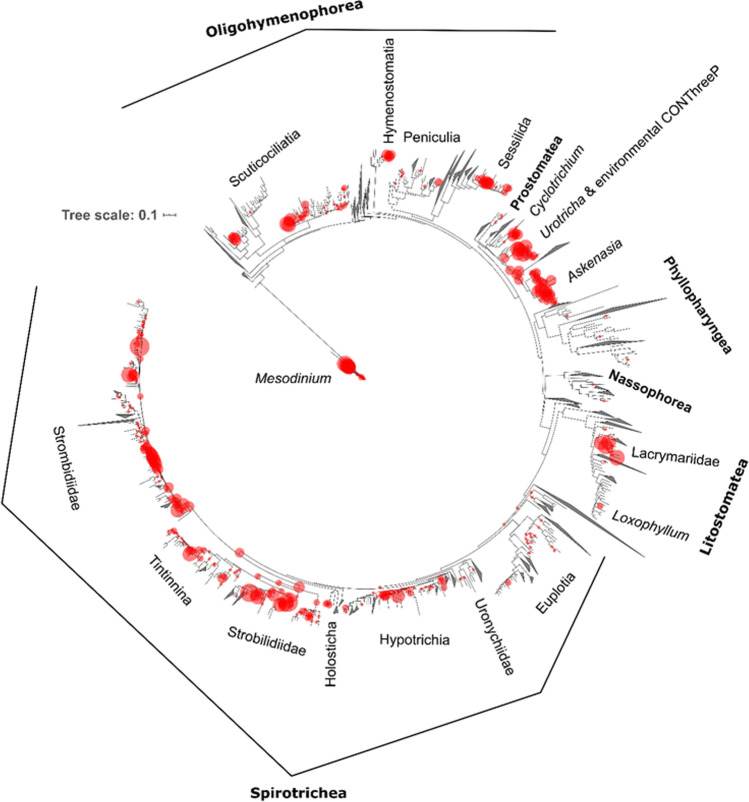
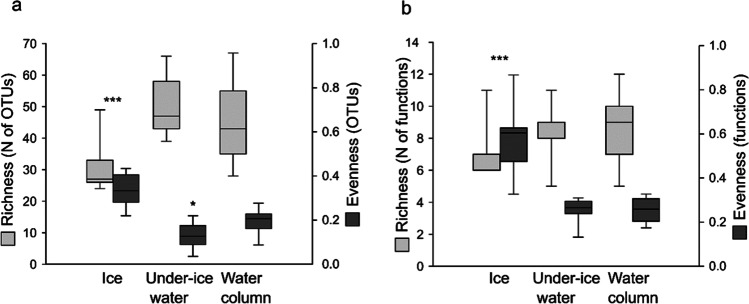
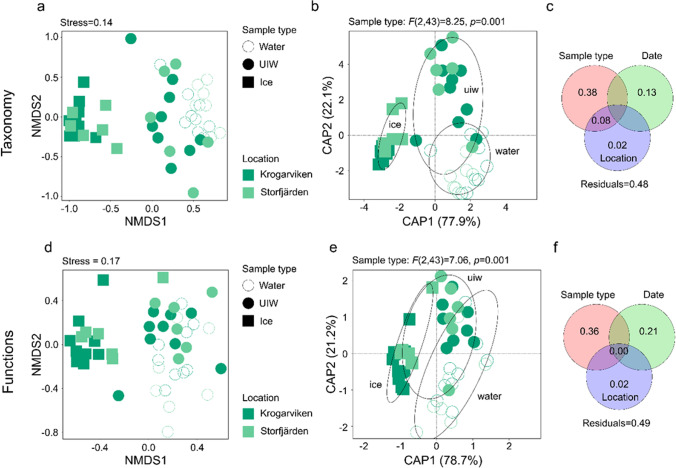
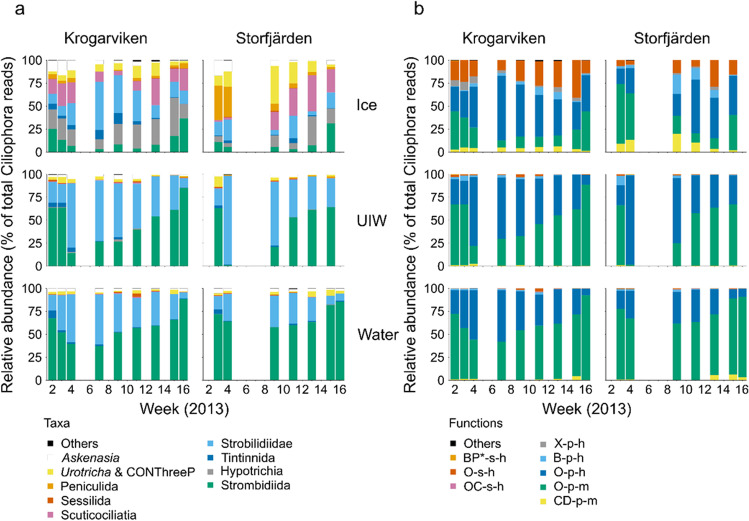
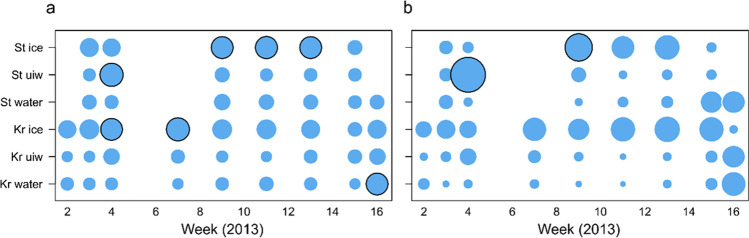
Ciliophora is a phylum of unicellular eukaryotes that are common and have pivotal roles in aquatic environments. Sea ice is a marine habitat, which is composed of a matrix of solid ice and pockets of saline water in which Ciliophora thrive. Here, we used phylogenetic placement to identify Ciliophora 18S ribosomal RNA reads obtained from wintertime water and sea ice, and assigned functions to the reads based on this taxonomic information. Based on our results, sea-ice Ciliophora assemblages are poorer in taxonomic and functional richness than under-ice water and water-column assemblages. Ciliophora diversity stayed stable throughout the ice-covered season both in sea ice and in water, although the assemblages changed during the course of our sampling. Under-ice water and the water column were distinctly predominated by planktonic orders Choreotrichida and Oligotrichida, which led to significantly lower taxonomic and functional evenness in water than in sea ice. In addition to planktonic Ciliophora, assemblages in sea ice included a set of moderately abundant surface-oriented species. Omnivory (feeding on bacteria and unicellular eukaryotes) was the most common feeding type but was not as predominant in sea ice as in water. Sea ice included cytotrophic (feeding on unicellular eukaryotes), bacterivorous and parasitic Ciliophora in addition to the predominant omnivorous Ciliophora. Potentially mixotrophic Ciliophora predominated the water column and heterotrophic Ciliophora sea ice. Our results highlight sea ice as an environment that creates a set of variable habitats, which may be threatened by the diminishing extent of sea ice due to changing climate.
Keywords: DNA metabarcoding; Mixotrophy; Phylogenetic placement; Predator–prey interactions; Winter ecology.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lynn DH. The ciliated protozoa. Characterization, classification, and guide to the literature. 3. New York: Springer; 2008.
-
- Adl SM, Bass D, Lane CE, Lukeš J, Schoch CL, Smirnov A, Agatha S, Berney C, Brown MW, Burki F, Cárdenas P, Čepička I, Chistyakova L, del Campo J, Dunthorn M, Edvardsen M, Eglit Y, Guillou L, Hampl V, Heiss AA, Hoppenrath M, James TY, Karnkowska A, Karpov S, Kim E, Kolisko M, Kudryavtsev A, Lahr DJG, Lara E, Le Gall L, Lynn DH, Mann DG, Massana R, Mitchell EAD, Morrow C, Park JS, Pawlowski JW, Powell MJ, Richter DJ, Rueckert S, Shadwick L, Shimano S, Spiegel FW, Torruella G, Youssef N, Zlatogursky V, Zhang Q. Revisions to the classification, nomenclature, and diversity of eukaryotes. J Eukaryot Microbiol. 2019;66:4–119. doi: 10.1111/jeu.12691. - DOI - PMC - PubMed
-
- Stoecker DK. Conceptual models of mixotrophy in planktonic protists and some ecological and evolutionary implications. Eur J Protistol. 1998;34:281–290. doi: 10.1016/S0932-4739(98)80055-2. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
