

Dynamic variability in SHP-1 abundance determines natural killer cell responsiveness
- PMID: 34752140
- PMCID: PMC8791782
- DOI: 10.1126/scisignal.abe5380
Dynamic variability in SHP-1 abundance determines natural killer cell responsiveness
Abstract
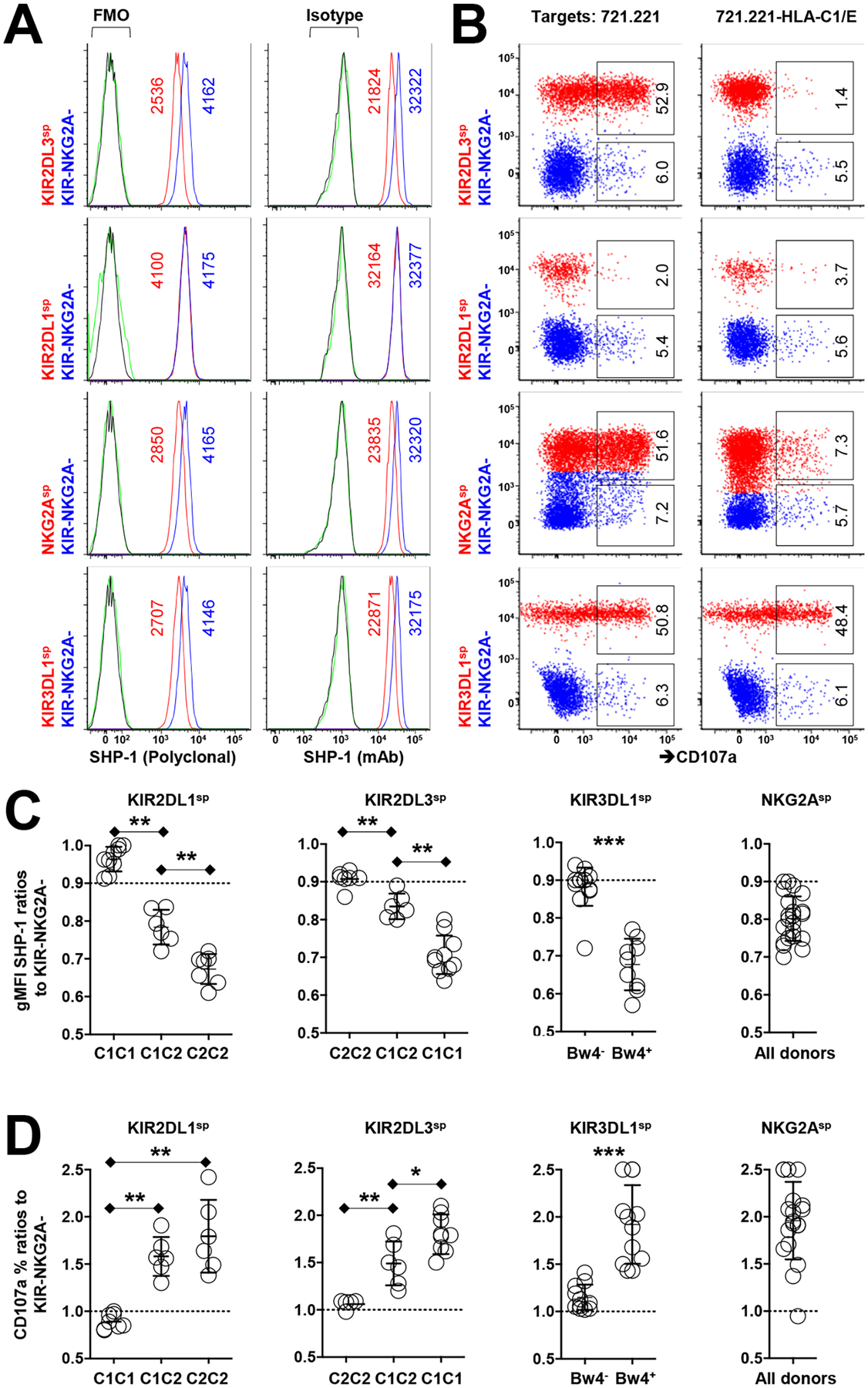
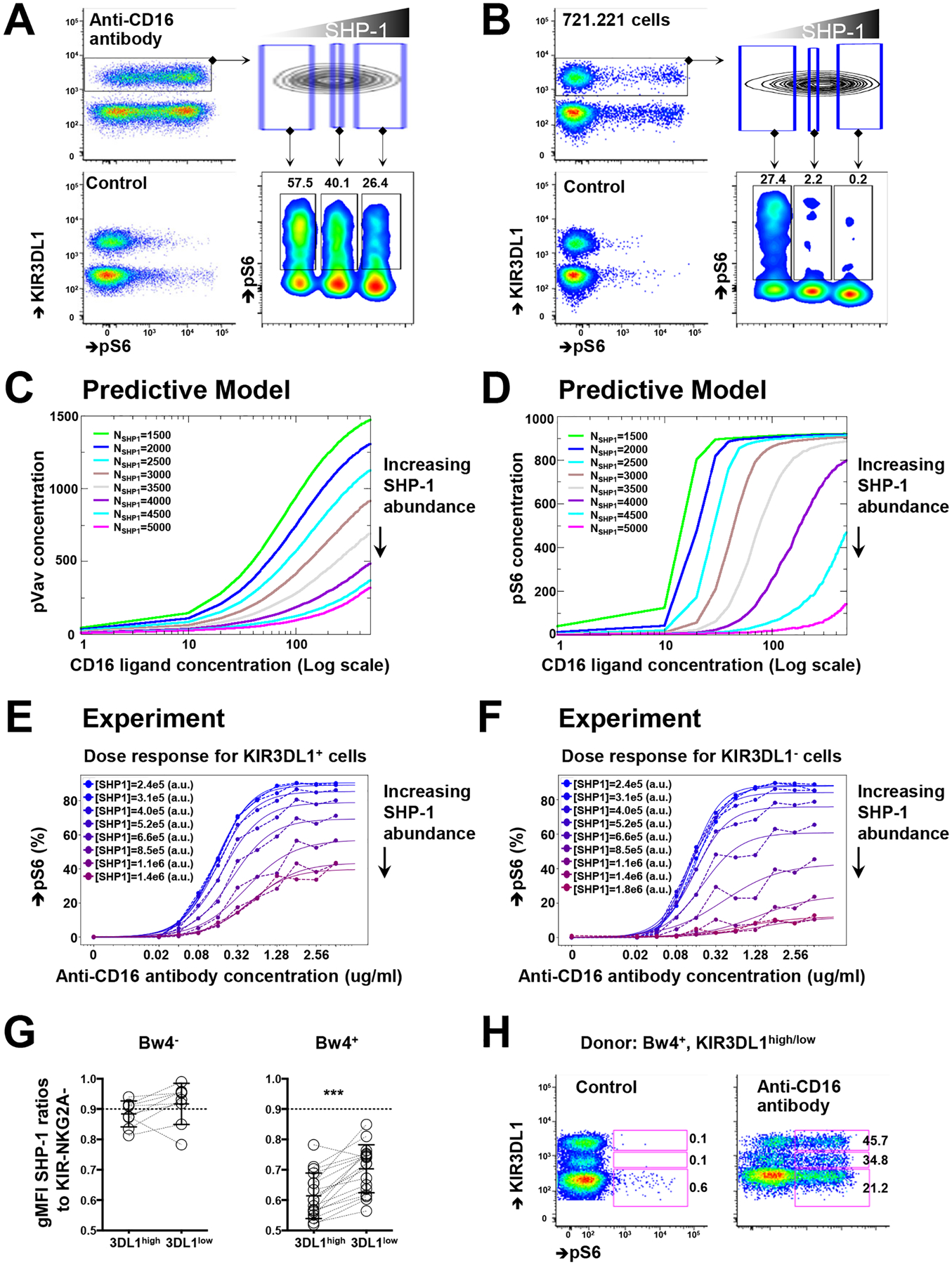
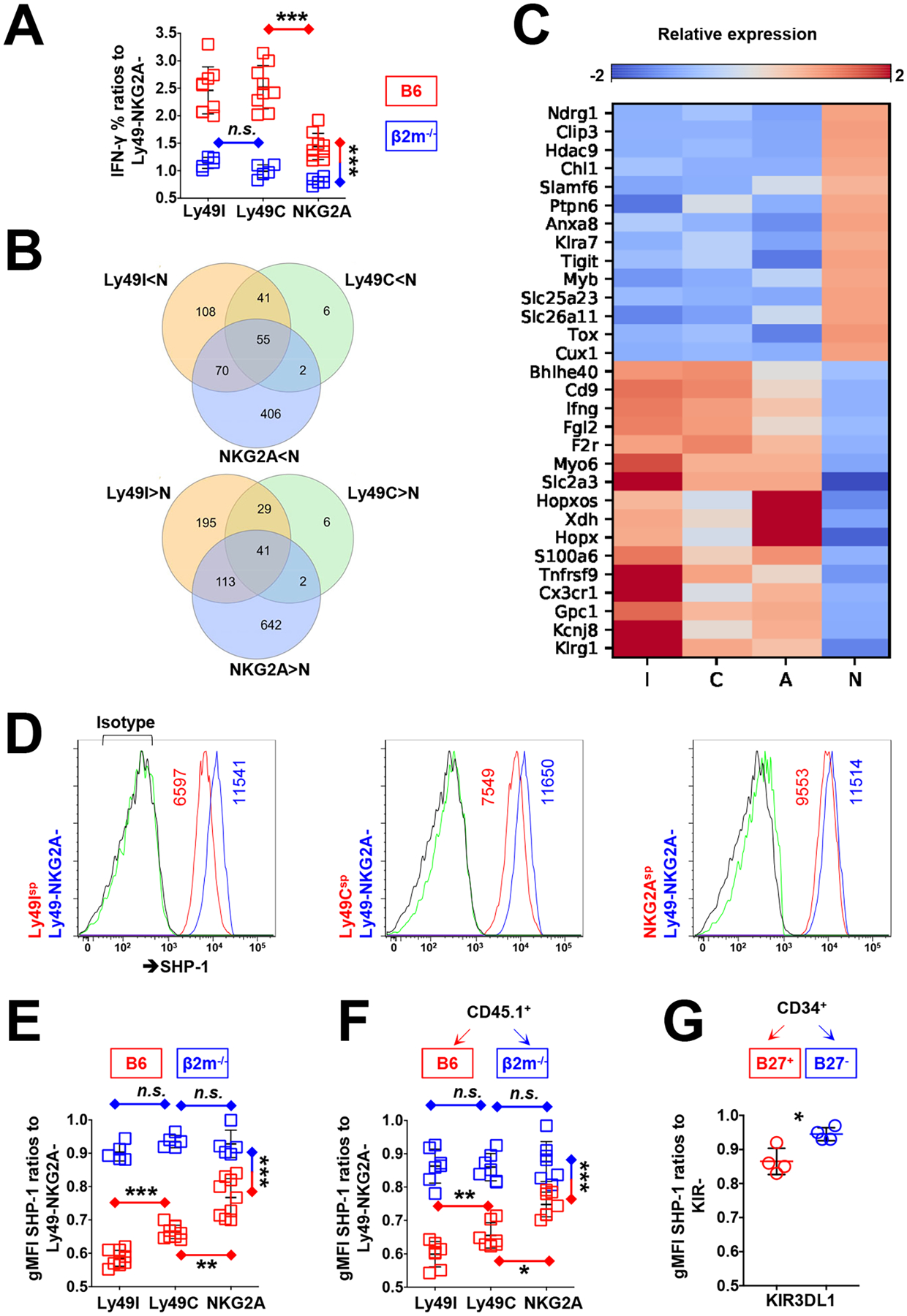
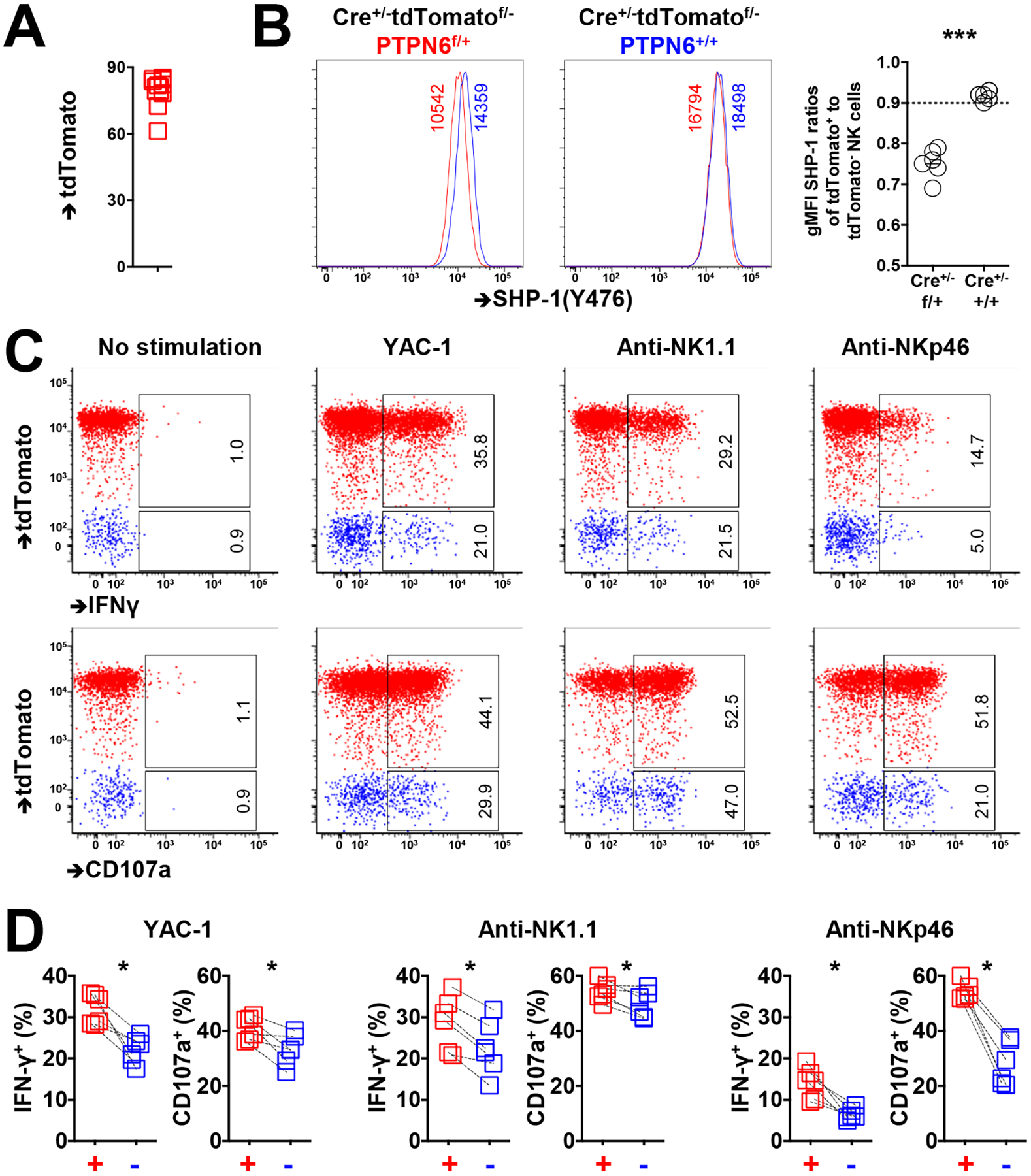
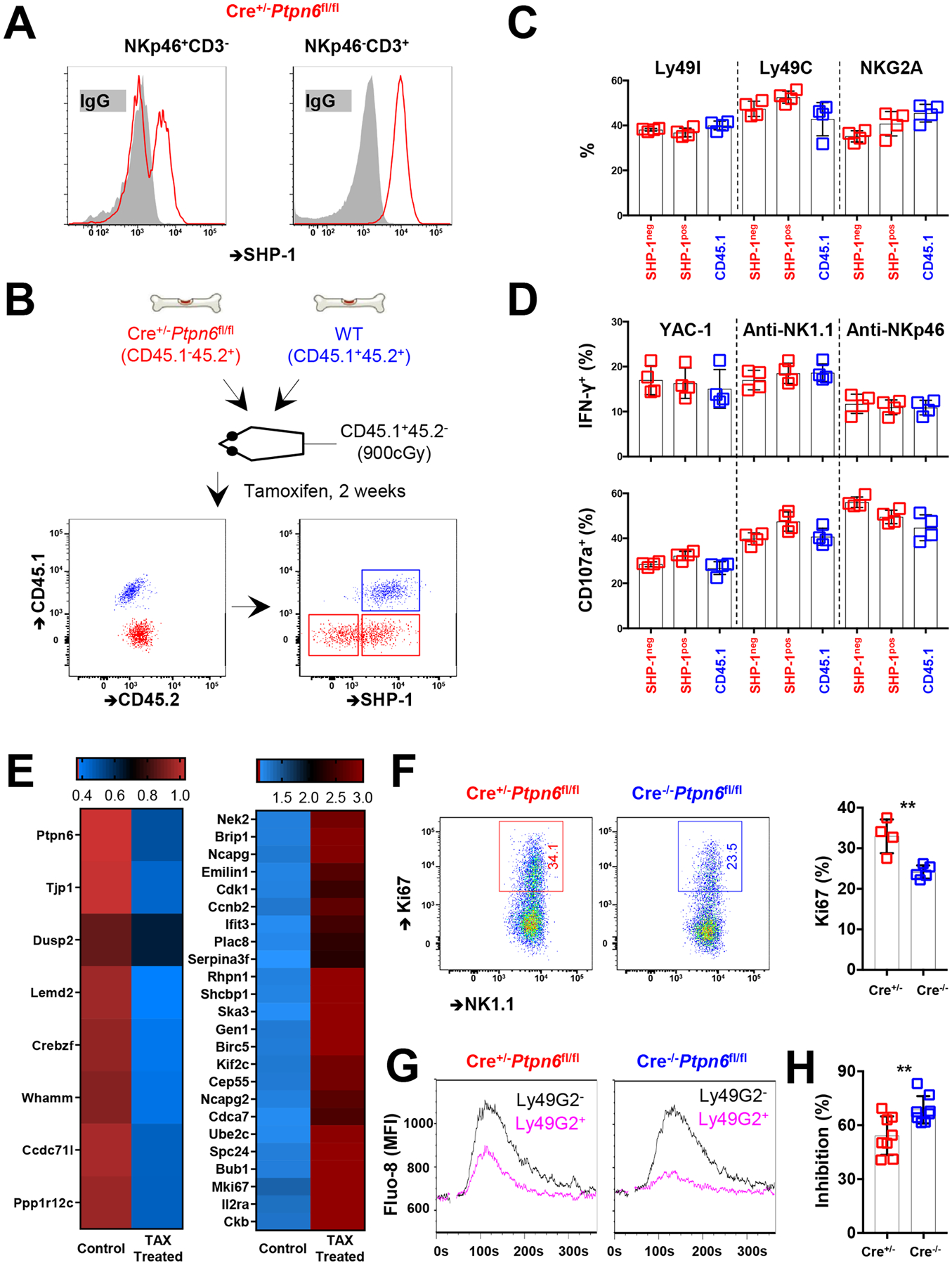
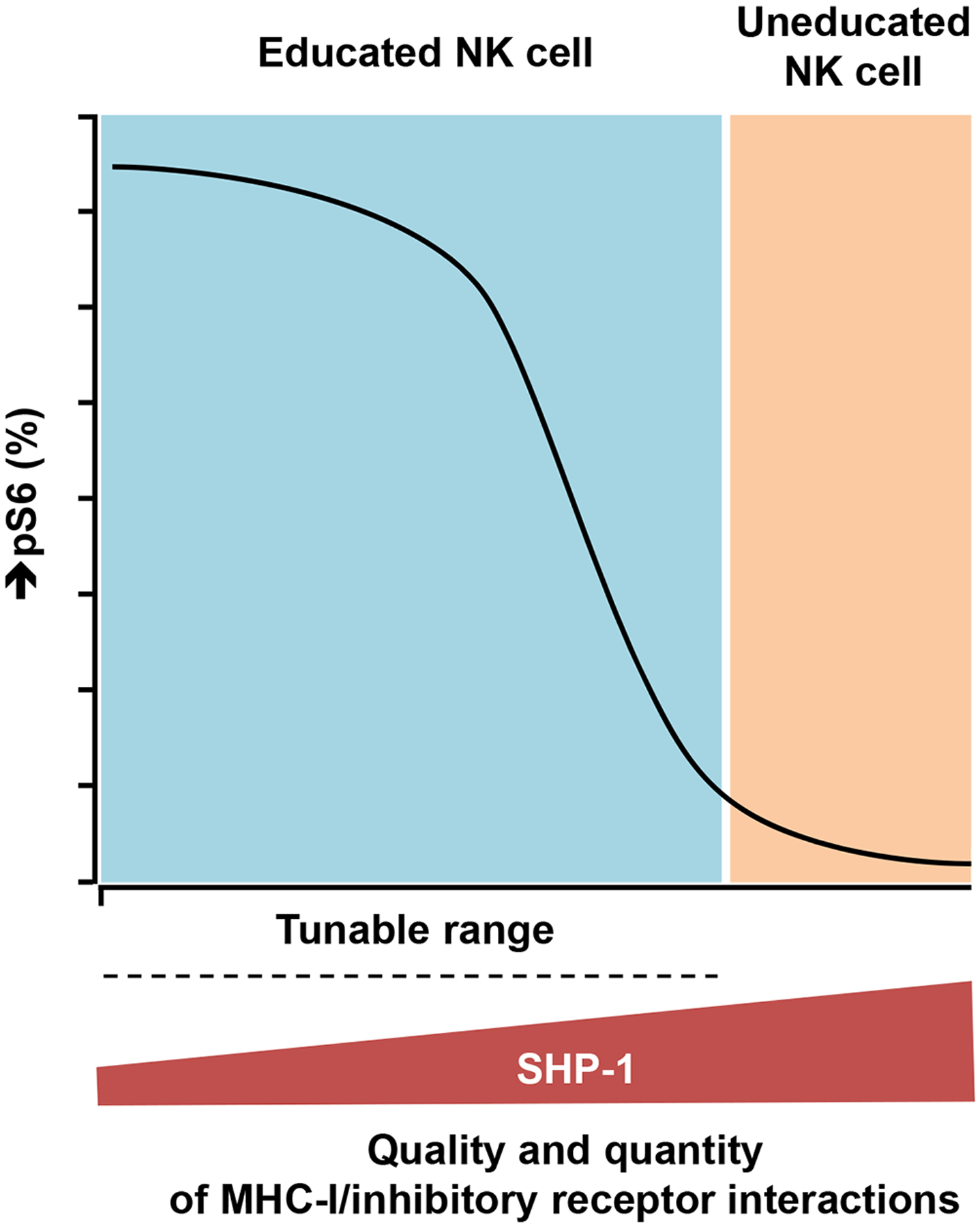
Interactions between human leukocyte antigen (HLA) molecules on target cells and the inhibitory killer cell immunoglobulin-like receptors (KIRs) and heterodimeric inhibitory receptor CD94-NKG2A on human natural killer (NK) cells shape and program various response capacities. A functionally orthologous system exists in mice, consisting of major histocompatibility complex (MHC) molecules on target cells and the inhibitory Ly49 and CD94-NKG2A receptors on NK cells. Here, we found that the abundance of Src homology 2 domain–containing phosphatase-1 (SHP-1) in NK cells was established by interactions between MHCs and NK cell inhibitory receptors, although phenotypically identical NK cell populations still showed substantial variability in endogenous SHP-1 abundance and NK cell response potential. Human and mouse NK cell populations with high responsiveness had low SHP-1 abundance, and a reduction in SHP-1 abundance in NK cells enhanced their responsiveness. Computational modeling of NK cell activation by membrane-proximal signaling events identified SHP-1 as a negative amplitude regulator, which was validated by single-cell analysis of human NK cell responsiveness. The amount of mRNA and protein varied among responsive NK cells despite their similar chromatin accessibility to that of unresponsive cells, suggesting dynamic regulation of SHP-1 abundance. Low intracellular SHP-1 abundance was a biomarker of responsive NK cells. Together, these data suggest that enhancing NK cell function through the acute loss of SHP-1 abundance or activity may enhance the tumoricidal capacity of NK cells.
Conflict of interest statement
Figures
References
-
- Lanier LL, NK cell recognition. Annu Rev Immunol 23, 225–274 (2005). - PubMed
-
- Karre K, Ljunggren HG, Piontek G, Kiessling R, Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature 319, 675–678 (1986). - PubMed
-
- Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, Yang L, French AR, Sunwoo JB, Lemieux S, Hansen TH, Yokoyama WM, Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature 436, 709–713 (2005). - PubMed
-
- Brodin P, Karre K, Hoglund P, NK cell education: not an on-off switch but a tunable rheostat. Trends Immunol 30, 143–149 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
