

CRISPR-Cas9 mediated mutation in GRAIN WIDTH and WEIGHT2 (GW2) locus improves aleurone layer and grain nutritional quality in rice
- PMID: 34753955
- PMCID: PMC8578329
- DOI: 10.1038/s41598-021-00828-z
CRISPR-Cas9 mediated mutation in GRAIN WIDTH and WEIGHT2 (GW2) locus improves aleurone layer and grain nutritional quality in rice
Abstract
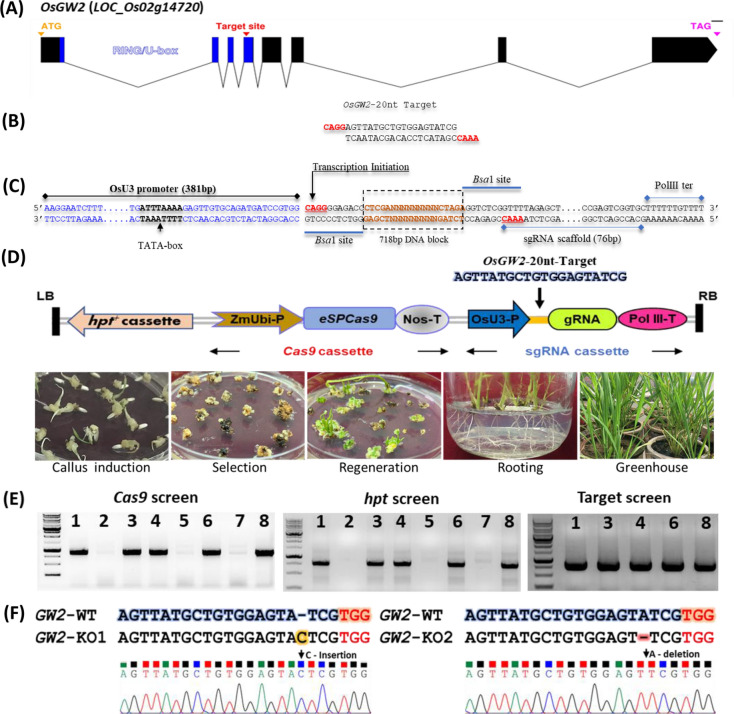
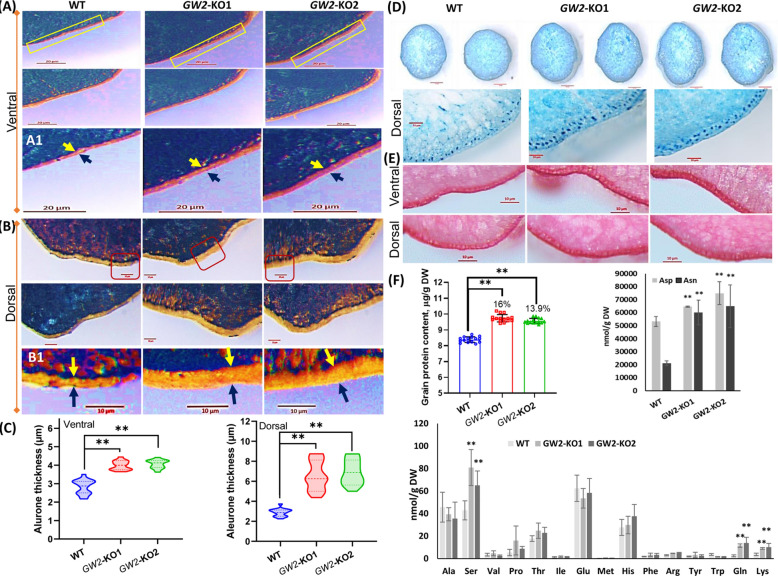
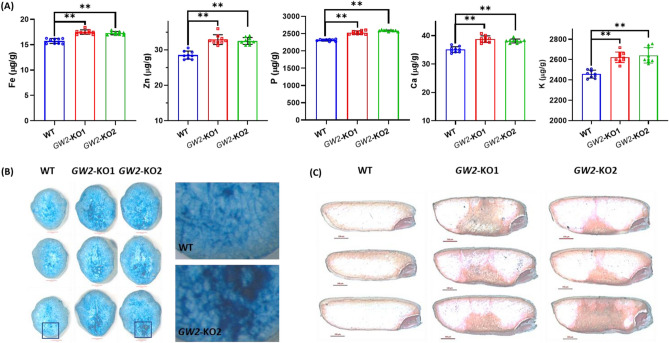
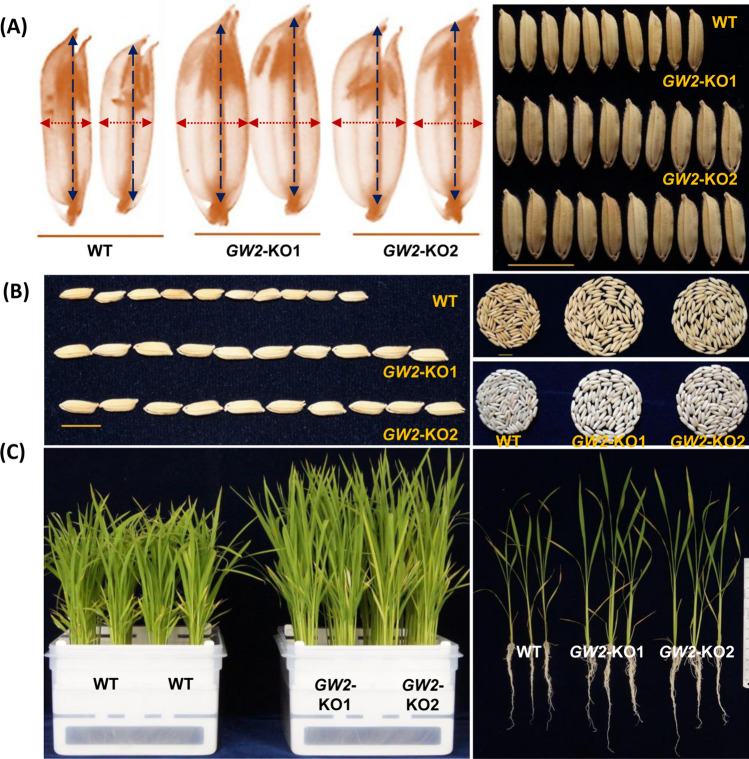
Enhancing crop productivity and their nutritional quality are the key components and primary focus of crop improvement strategy for fulfilling future food demand and improving human health. Grain filling and endosperm development are the key determinants of grain yield and nutritional quality. GRAIN WIDTH and WEIGHT2 (GW2) gene encodes a RING-type E3 ubiquitin ligase and determines the grain weight in cereal crops. Here we report GW2 knockout (KO) mutants in Indica (var. MTU1010) through CRISPR/Cas9 genome editing. The endosperm of GW2-KO mutant seed displays a thick aleurone layer with enhanced grain protein content. Further the loss of function of OsGW2 results in improved accumulation of essential dietary minerals (Fe, Zn, K, P, Ca) in the endosperm of rice grain. Additionally, the mutants displayed an early growth vigour phenotype with an improved root and shoot architecture. The hull morphology of GW2-KO lines also showed improved, grain filling thereby promoting larger grain architecture. Together, our findings indicate that GW2 may serve as a key regulator of improved grain architecture, grain nutritional quality and an important modulator of plant morphology. The study offers a strategy for the development of improved rice cultivars with enriched nutritional quality and its possible implementation in other cereals as well.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
