

Phase transition specified by a binary code patterns the vertebrate eye cup
- PMID: 34757798
- PMCID: PMC8580326
- DOI: 10.1126/sciadv.abj9846
Phase transition specified by a binary code patterns the vertebrate eye cup
Abstract
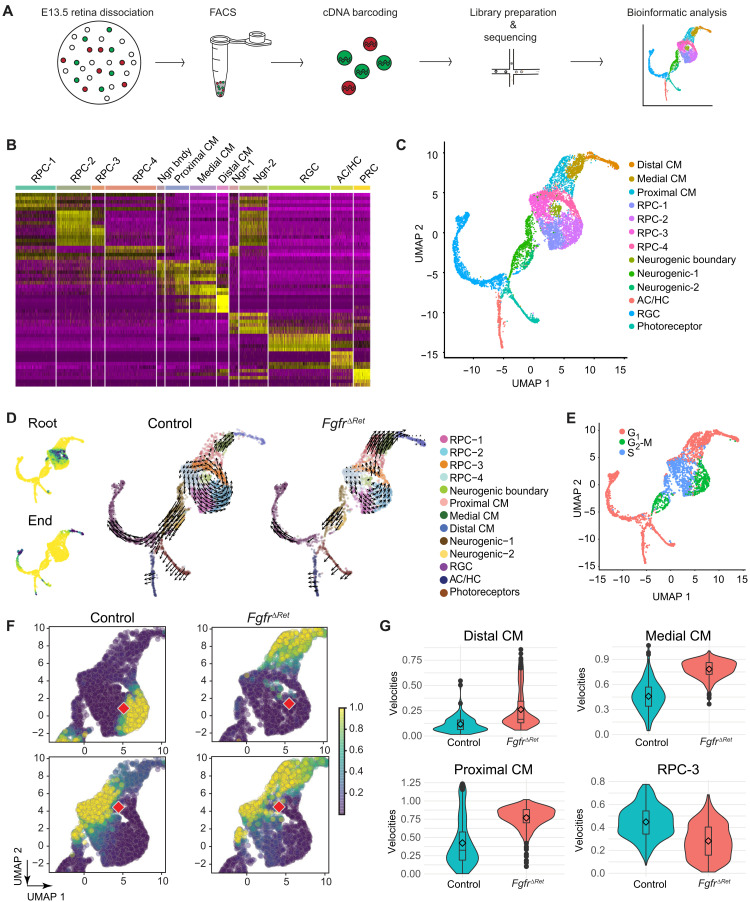
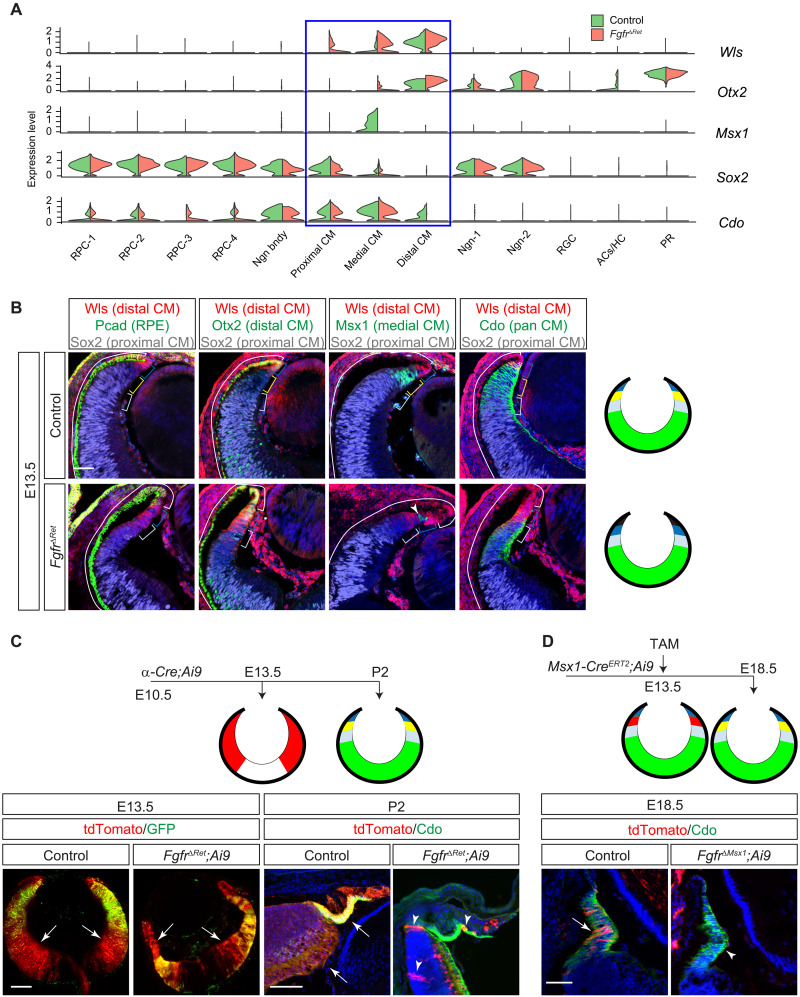
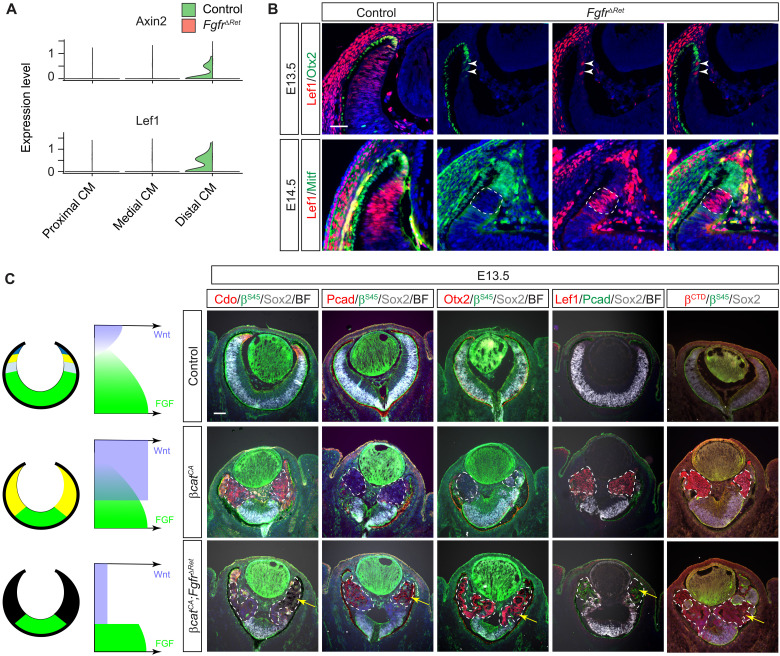
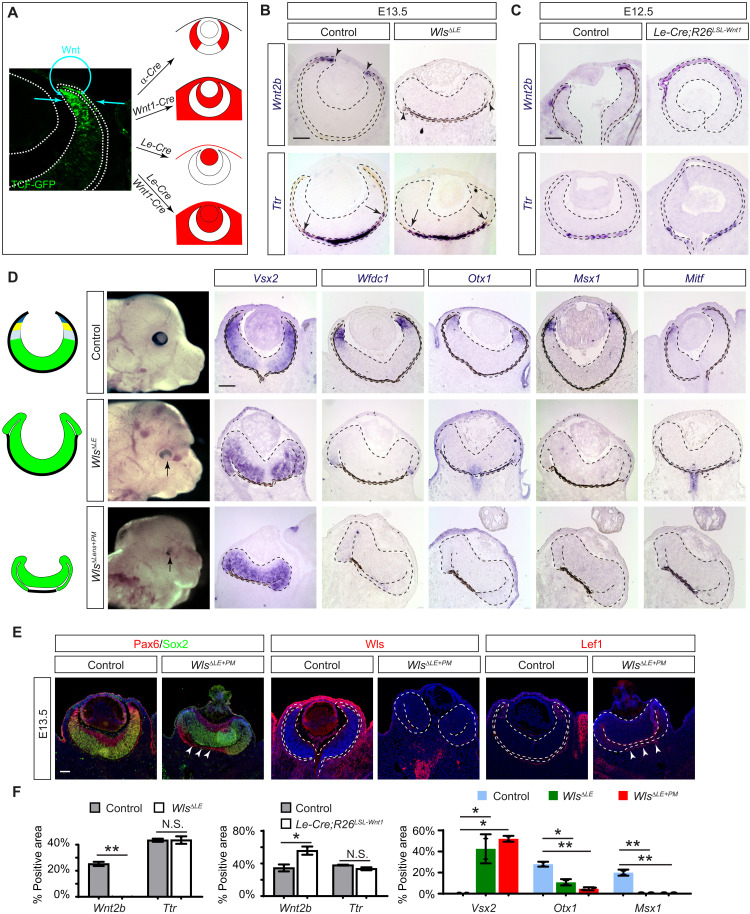
The developing vertebrate eye cup is partitioned into the neural retina (NR), the retinal pigmented epithelium (RPE), and the ciliary margin (CM). By single-cell analysis, we showed that fibroblast growth factor (FGF) signaling regulates the CM in its stem cell–like property of self-renewal, differentiation, and survival, which is balanced by an evolutionarily conserved Wnt signaling gradient. FGF promotes Wnt signaling in the CM by stabilizing β-catenin in a GSK3β-independent manner. While Wnt signaling converts the NR to either the CM or the RPE depending on FGF signaling, FGF transforms the RPE to the NR or CM dependent on Wnt activity. The default fate of the eye cup is the NR, but synergistic FGF and Wnt signaling promotes CM formation both in vivo and in human retinal organoid. Our study reveals that the vertebrate eye develops through phase transition determined by a combinatorial code of FGF and Wnt signaling.
Figures



References
-
- Chow R. L., Lang R. A., Early eye development in vertebrates. Annu. Rev. Cell Dev. Biol. 17, 255–296 (2001). - PubMed
-
- Tomlinson A., Patterning the peripheral retina of the fly: Decoding a gradient. Dev. Cell 5, 799–809 (2003). - PubMed
-
- Fujimura N., Taketo M. M., Mori M., Korinek V., Kozmik Z., Spatial and temporal regulation of Wnt/β-catenin signaling is essential for development of the retinal pigment epithelium. Dev. Biol. 334, 31–45 (2009). - PubMed
Grants and funding
- R21 AG050437/AG/NIA NIH HHS/United States
- R01 EY031354/EY/NEI NIH HHS/United States
- R01 EY017061/EY/NEI NIH HHS/United States
- R01 EY018213/EY/NEI NIH HHS/United States
- U01 EY030580/EY/NEI NIH HHS/United States
- U54 OD020351/OD/NIH HHS/United States
- R01 EY026682/EY/NEI NIH HHS/United States
- R01 EY025933/EY/NEI NIH HHS/United States
- R01 HD106120/HD/NICHD NIH HHS/United States
- R24 EY028758/EY/NEI NIH HHS/United States
- R01 EY024698/EY/NEI NIH HHS/United States
- P30 EY019007/EY/NEI NIH HHS/United States
- R24 EY027285/EY/NEI NIH HHS/United States
- R01 DK113642/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
