

A single amino acid polymorphism in a conserved effector of the multihost blast fungus pathogen expands host-target binding spectrum
- PMID: 34758051
- PMCID: PMC8608293
- DOI: 10.1371/journal.ppat.1009957
A single amino acid polymorphism in a conserved effector of the multihost blast fungus pathogen expands host-target binding spectrum
Abstract
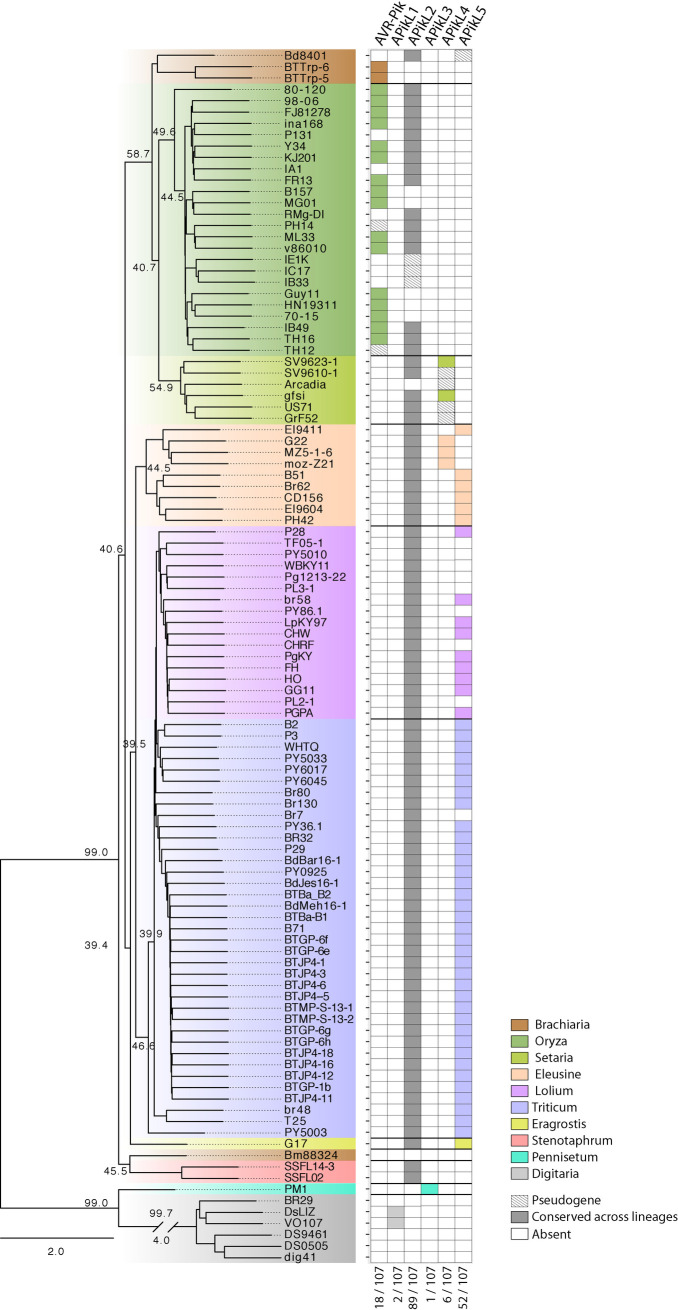
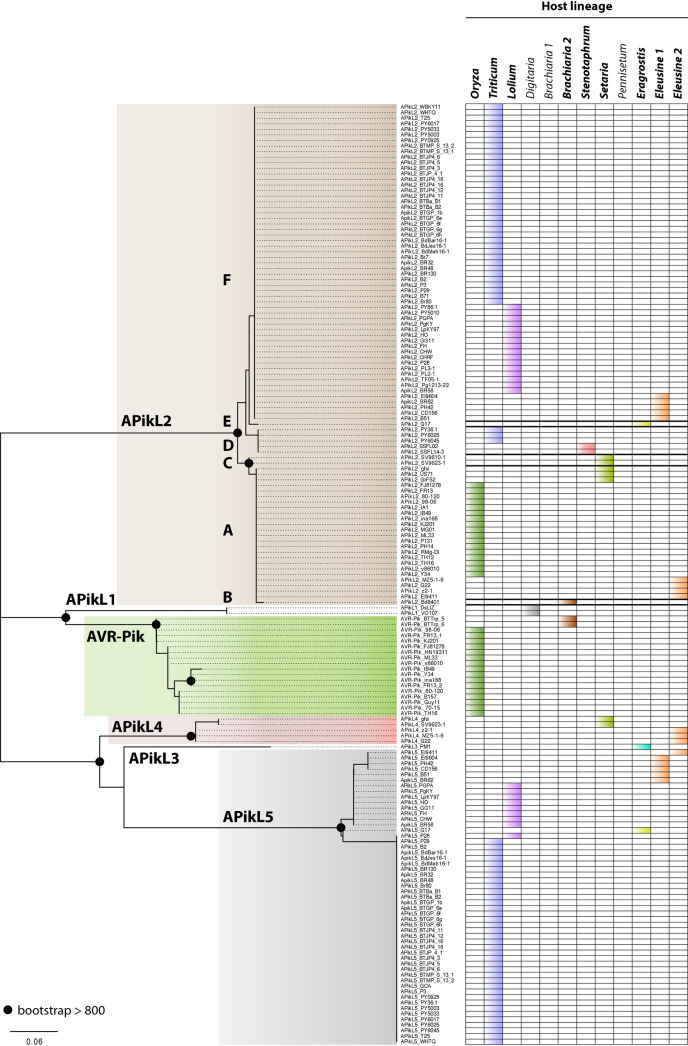
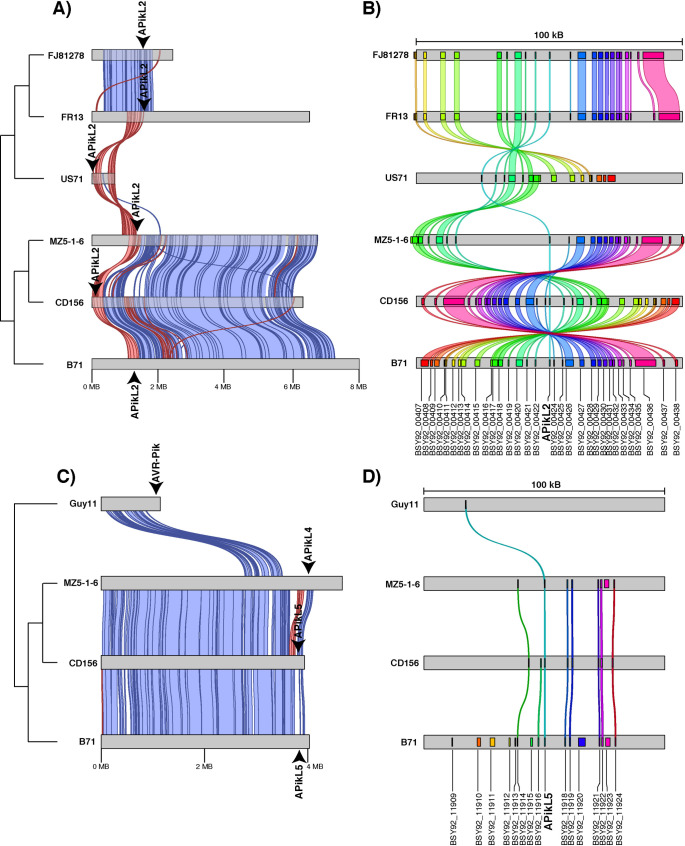
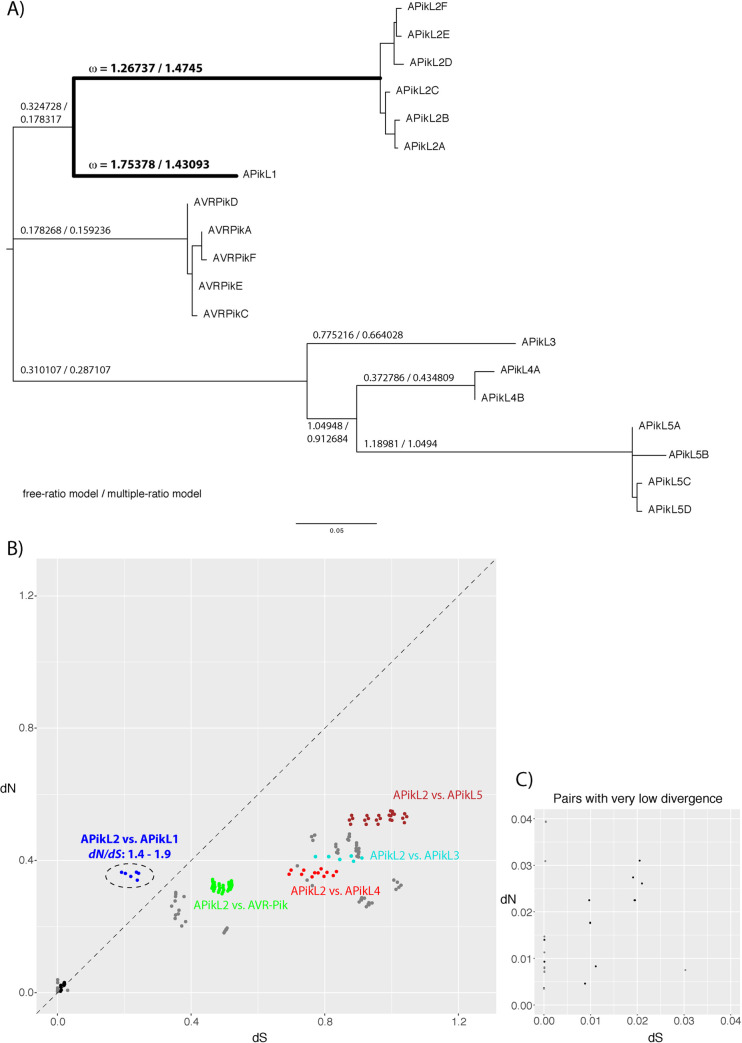
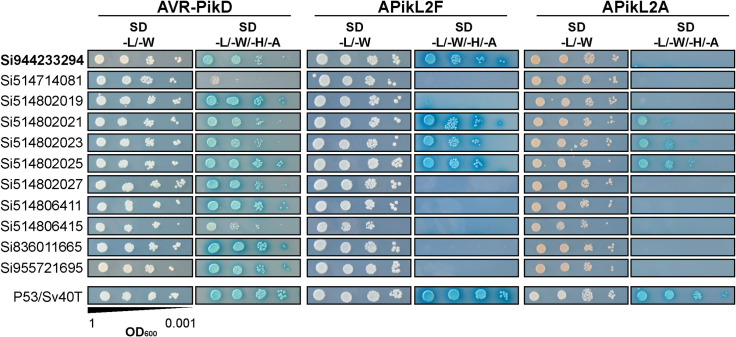
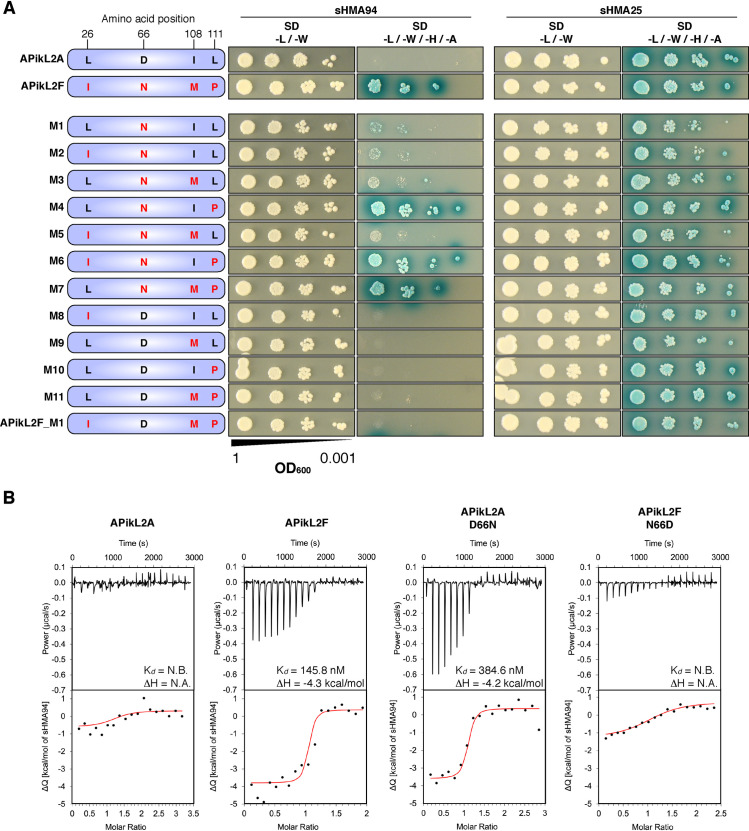
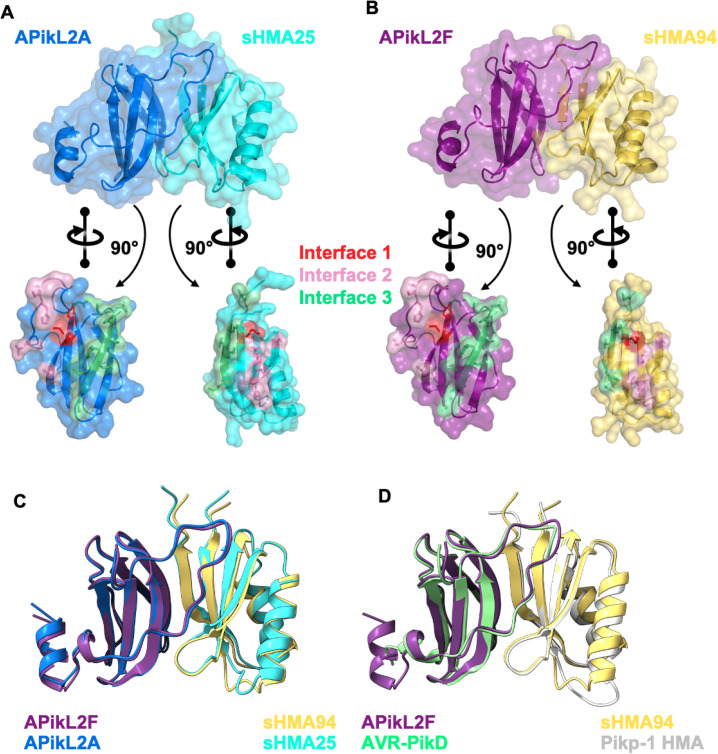
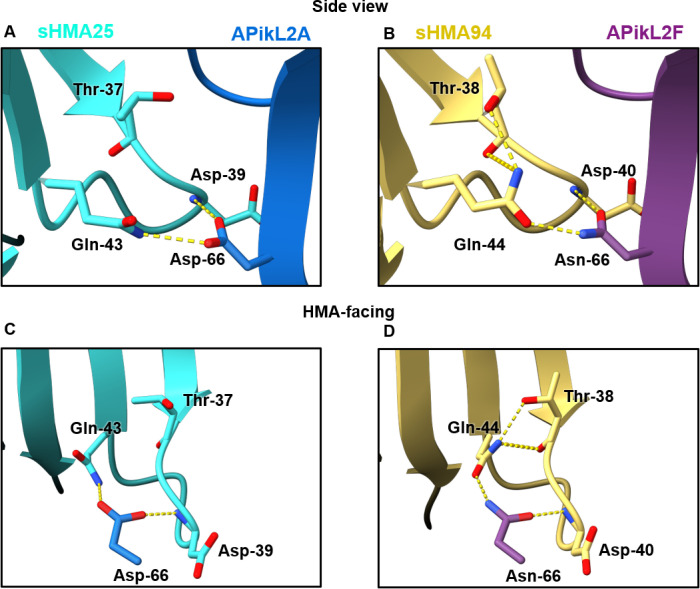
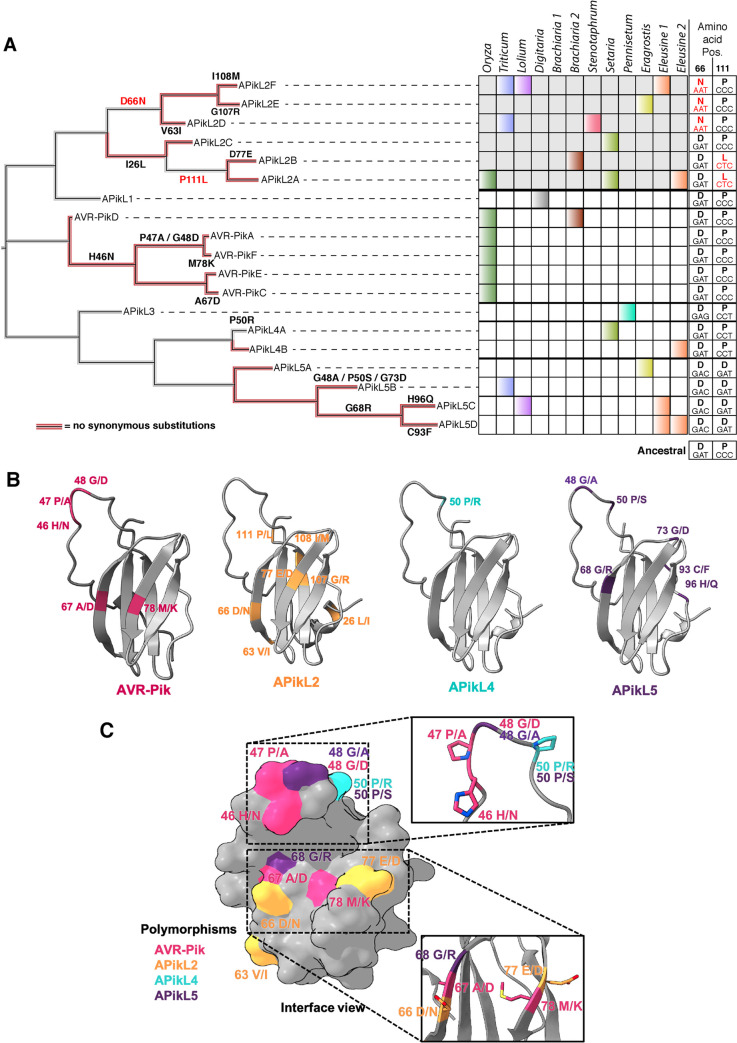
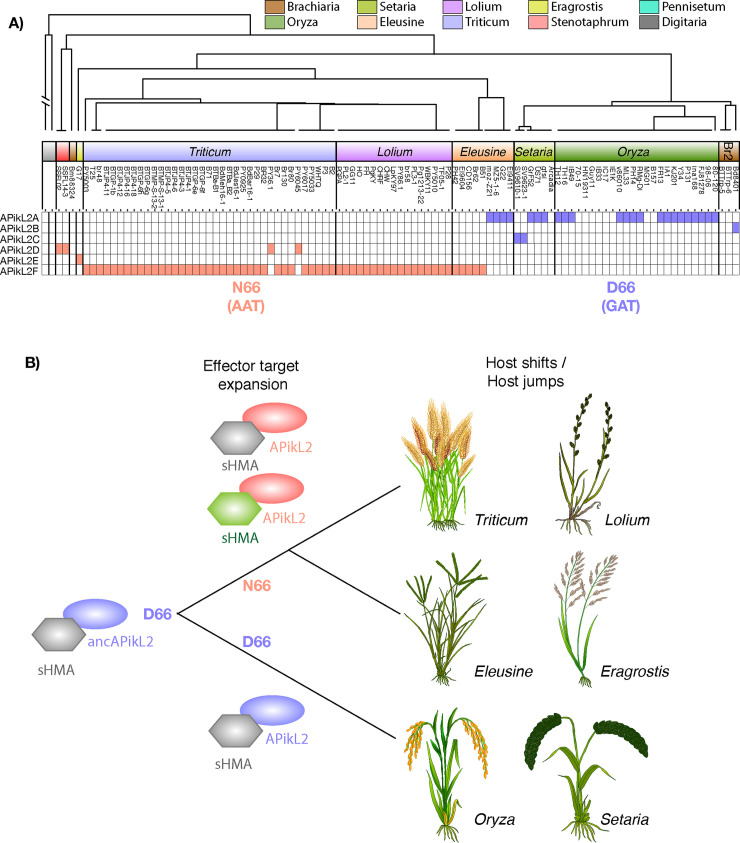
Accelerated gene evolution is a hallmark of pathogen adaptation and specialization following host-jumps. However, the molecular processes associated with adaptive evolution between host-specific lineages of a multihost plant pathogen remain poorly understood. In the blast fungus Magnaporthe oryzae (Syn. Pyricularia oryzae), host specialization on different grass hosts is generally associated with dynamic patterns of gain and loss of virulence effector genes that tend to define the distinct genetic lineages of this pathogen. Here, we unravelled the biochemical and structural basis of adaptive evolution of APikL2, an exceptionally conserved paralog of the well-studied rice-lineage specific effector AVR-Pik. Whereas AVR-Pik and other members of the six-gene AVR-Pik family show specific patterns of presence/absence polymorphisms between grass-specific lineages of M. oryzae, APikL2 stands out by being ubiquitously present in all blast fungus lineages from 13 different host species. Using biochemical, biophysical and structural biology methods, we show that a single aspartate to asparagine polymorphism expands the binding spectrum of APikL2 to host proteins of the heavy-metal associated (HMA) domain family. This mutation maps to one of the APikL2-HMA binding interfaces and contributes to an altered hydrogen-bonding network. By combining phylogenetic ancestral reconstruction with an analysis of the structural consequences of allelic diversification, we revealed a common mechanism of effector specialization in the AVR-Pik/APikL2 family that involves two major HMA-binding interfaces. Together, our findings provide a detailed molecular evolution and structural biology framework for diversification and adaptation of a fungal pathogen effector family following host-jumps.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Multiple variants of the fungal effector AVR-Pik bind the HMA domain of the rice protein OsHIPP19, providing a foundation to engineer plant defense.J Biol Chem. 2021 Jan-Jun;296:100371. doi: 10.1016/j.jbc.2021.100371. Epub 2021 Feb 4. J Biol Chem. 2021. PMID: 33548226 Free PMC article.
-
The allelic rice immune receptor Pikh confers extended resistance to strains of the blast fungus through a single polymorphism in the effector binding interface.PLoS Pathog. 2021 Mar 1;17(3):e1009368. doi: 10.1371/journal.ppat.1009368. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33647072 Free PMC article.
-
Polymorphic residues in rice NLRs expand binding and response to effectors of the blast pathogen.Nat Plants. 2018 Aug;4(8):576-585. doi: 10.1038/s41477-018-0194-x. Epub 2018 Jul 9. Nat Plants. 2018. PMID: 29988155
-
Investigating the cell biology of plant infection by the rice blast fungus Magnaporthe oryzae.Curr Opin Microbiol. 2016 Dec;34:147-153. doi: 10.1016/j.mib.2016.10.001. Epub 2016 Nov 3. Curr Opin Microbiol. 2016. PMID: 27816794 Review.
-
Dynamic Gene-for-Gene Interactions Undermine Durable Resistance.Mol Plant Microbe Interact. 2025 Mar;38(2):104-117. doi: 10.1094/MPMI-02-25-0022-HH. Epub 2025 Apr 24. Mol Plant Microbe Interact. 2025. PMID: 40272515 Review.
Cited by
-
Bioengineering a plant NLR immune receptor with a robust binding interface toward a conserved fungal pathogen effector.Proc Natl Acad Sci U S A. 2024 Jul 9;121(28):e2402872121. doi: 10.1073/pnas.2402872121. Epub 2024 Jul 5. Proc Natl Acad Sci U S A. 2024. PMID: 38968126 Free PMC article.
-
Two NLR immune receptors acquired high-affinity binding to a fungal effector through convergent evolution of their integrated domain.Elife. 2021 Jul 21;10:e66961. doi: 10.7554/eLife.66961. Elife. 2021. PMID: 34288868 Free PMC article.
-
Functional diversification gave rise to allelic specialization in a rice NLR immune receptor pair.Elife. 2021 Nov 16;10:e71662. doi: 10.7554/eLife.71662. Elife. 2021. PMID: 34783652 Free PMC article.
-
Does a Similar 3D Structure Mean a Similar Folding Pathway? The Presence of a C-Terminal α-Helical Extension in the 3D Structure of MAX60 Drastically Changes the Folding Pathway Described for Other MAX-Effectors from Magnaporthe oryzae.Molecules. 2023 Aug 15;28(16):6068. doi: 10.3390/molecules28166068. Molecules. 2023. PMID: 37630320 Free PMC article.
-
Exploiting Structural Modelling Tools to Explore Host-Translocated Effector Proteins.Int J Mol Sci. 2021 Nov 30;22(23):12962. doi: 10.3390/ijms222312962. Int J Mol Sci. 2021. PMID: 34884778 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
