

Host immunomodulatory lipids created by symbionts from dietary amino acids
- PMID: 34759313
- PMCID: PMC8999822
- DOI: 10.1038/s41586-021-04083-0
Host immunomodulatory lipids created by symbionts from dietary amino acids
Erratum in
-
Publisher Correction: Host immunomodulatory lipids created by symbionts from dietary amino acids.Nature. 2022 Jan;601(7894):E32. doi: 10.1038/s41586-021-04276-7. Nature. 2022. PMID: 35001086 No abstract available.
Abstract
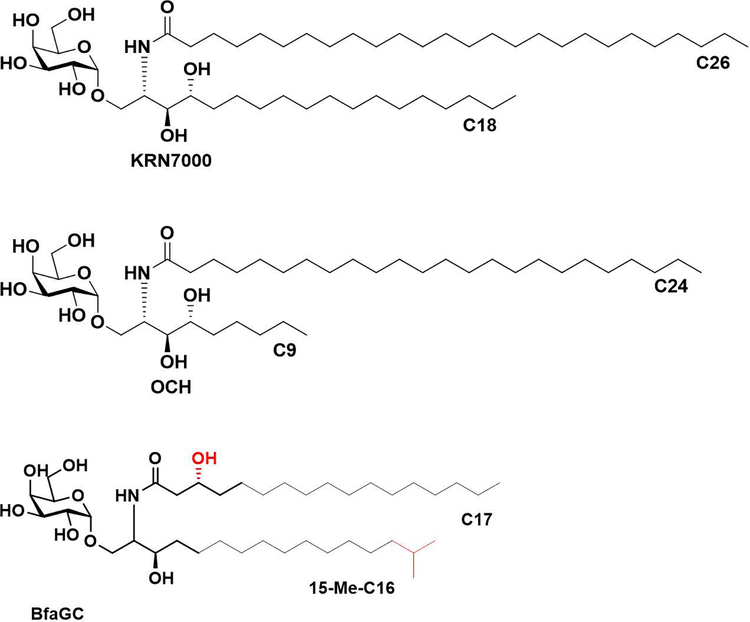
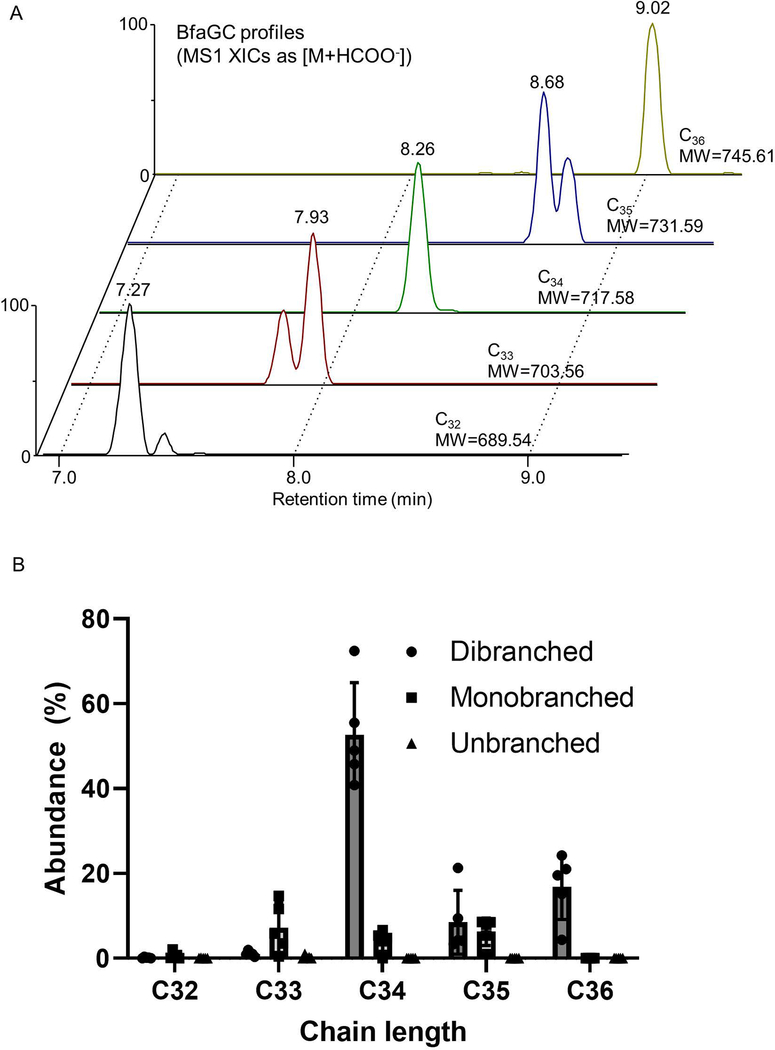
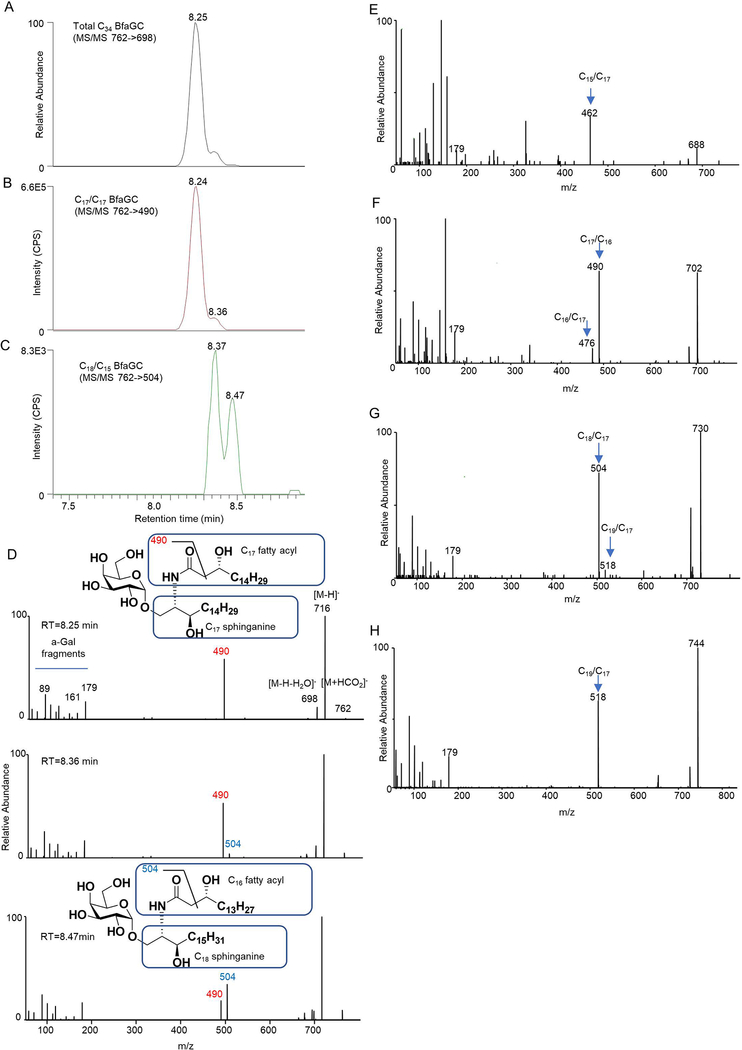
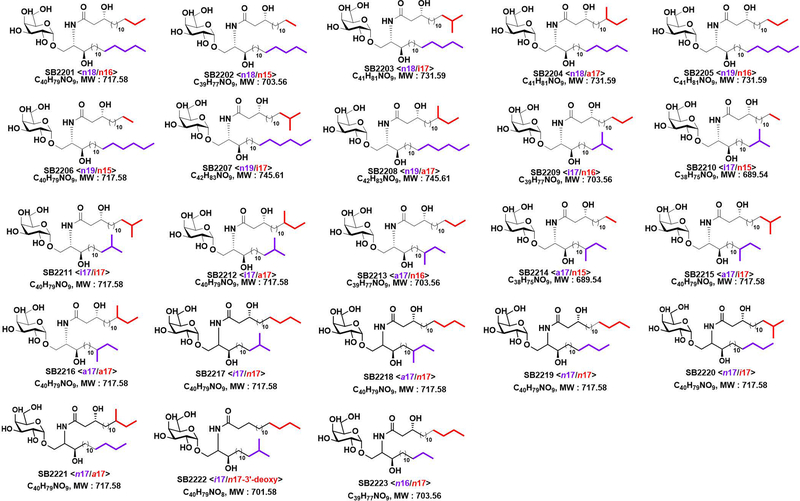
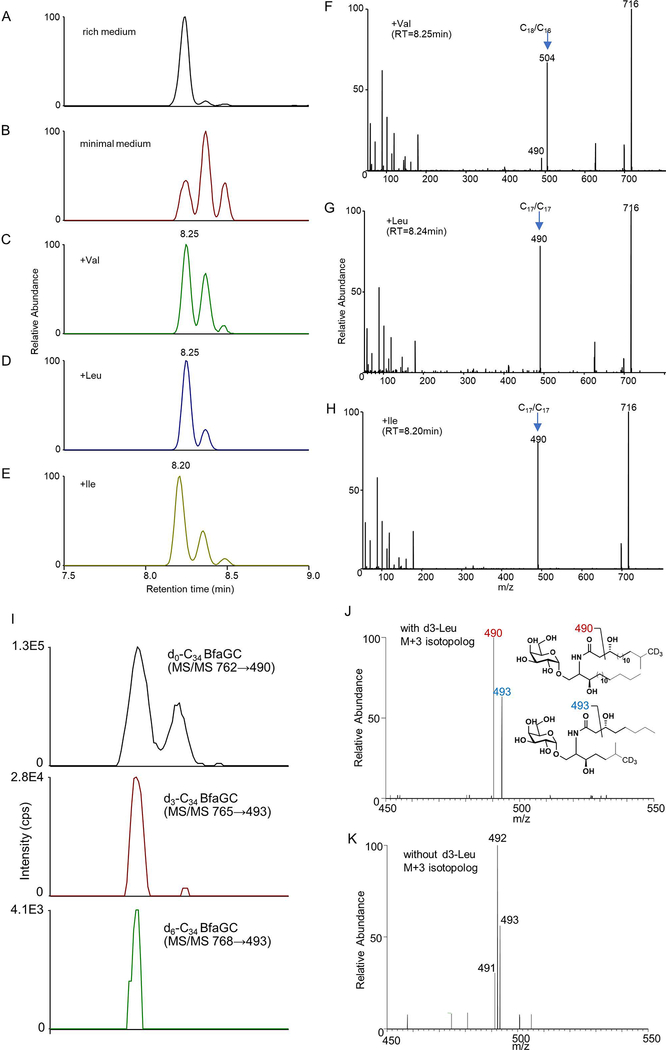
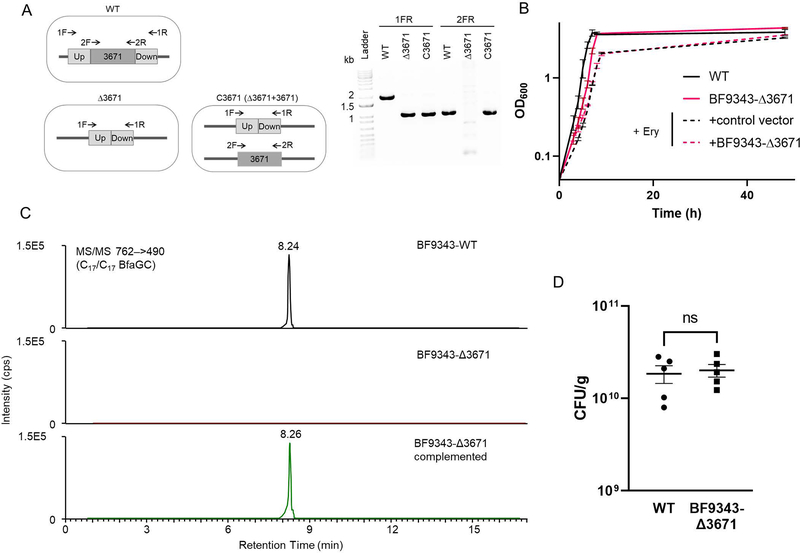
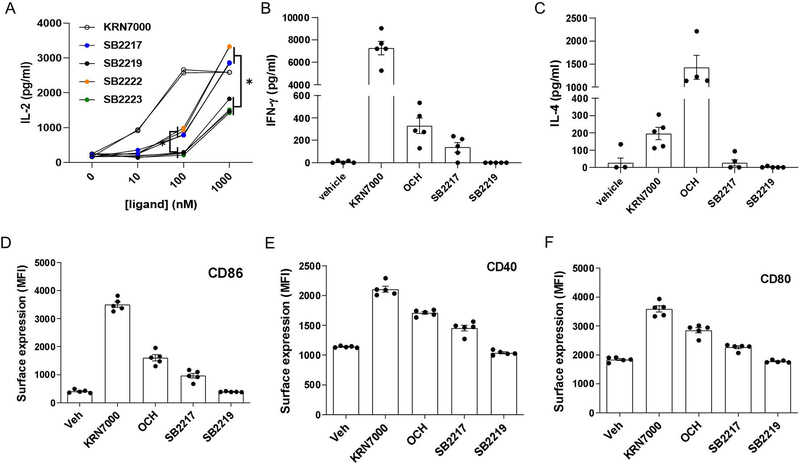
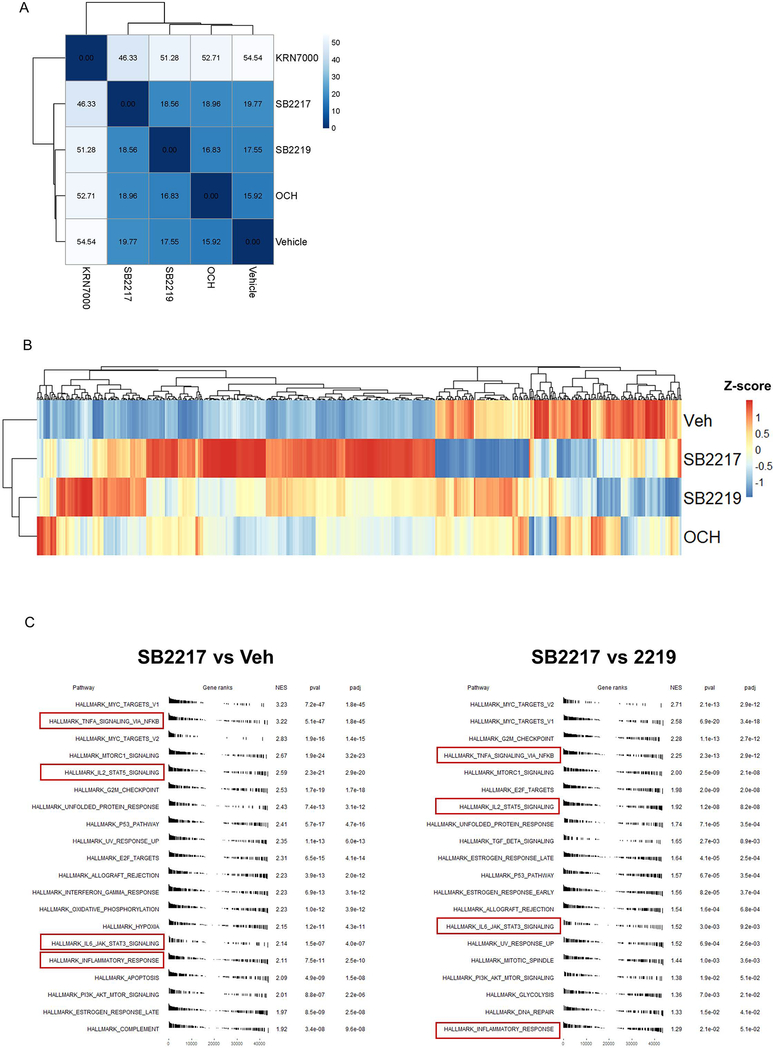
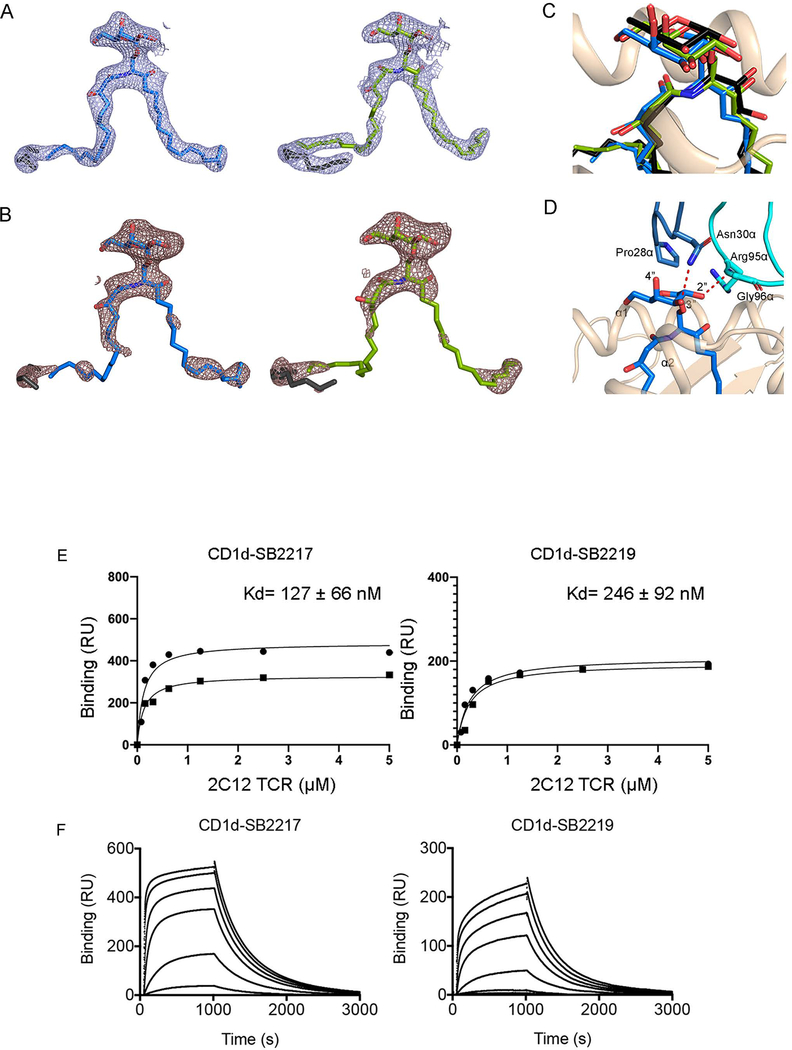
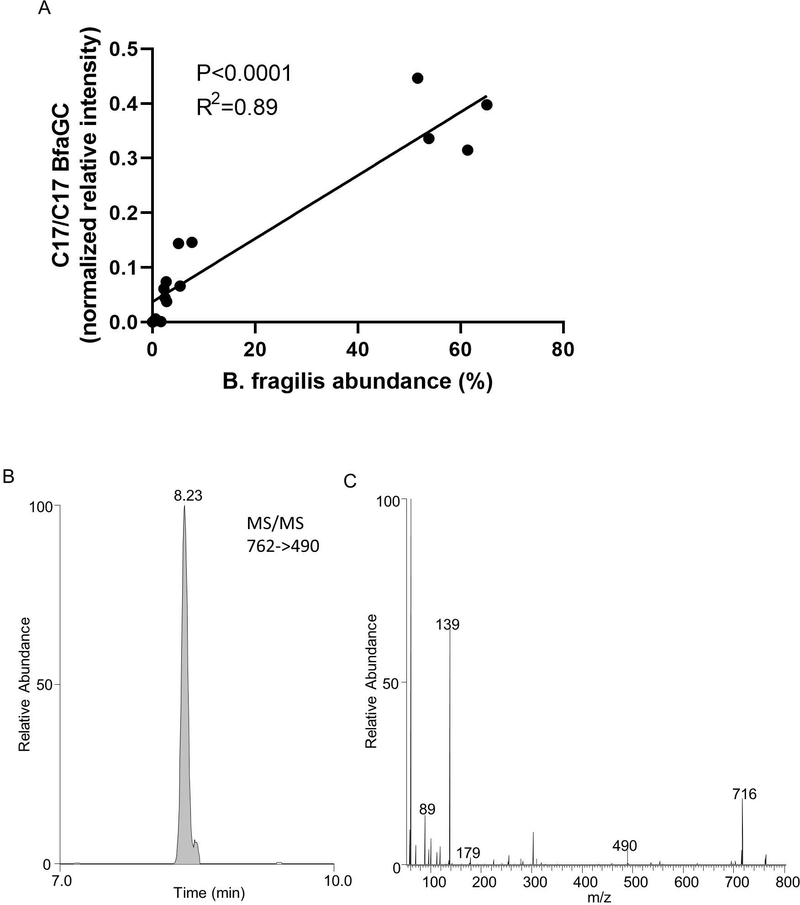
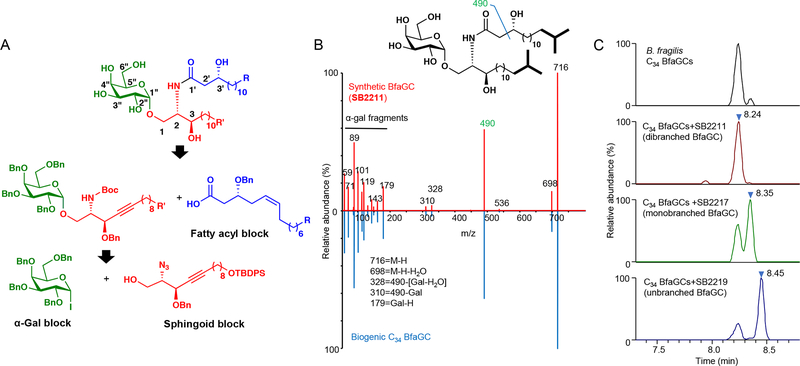
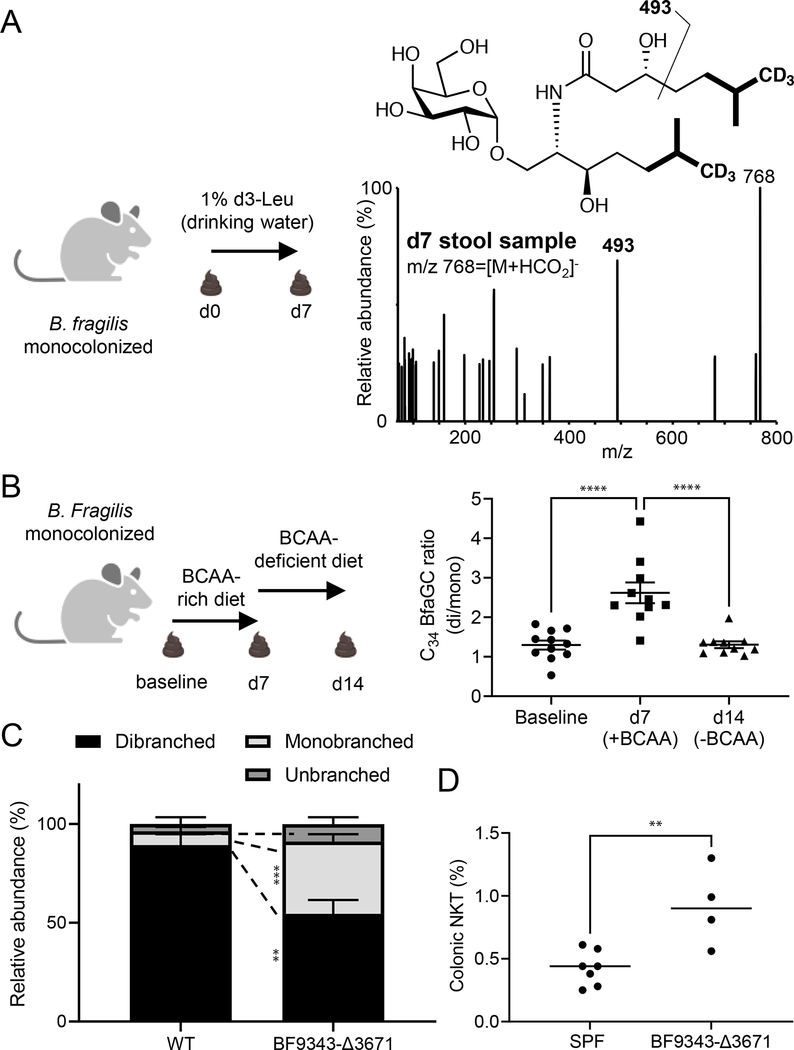
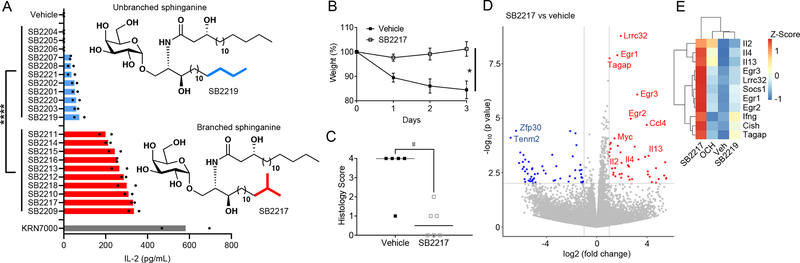
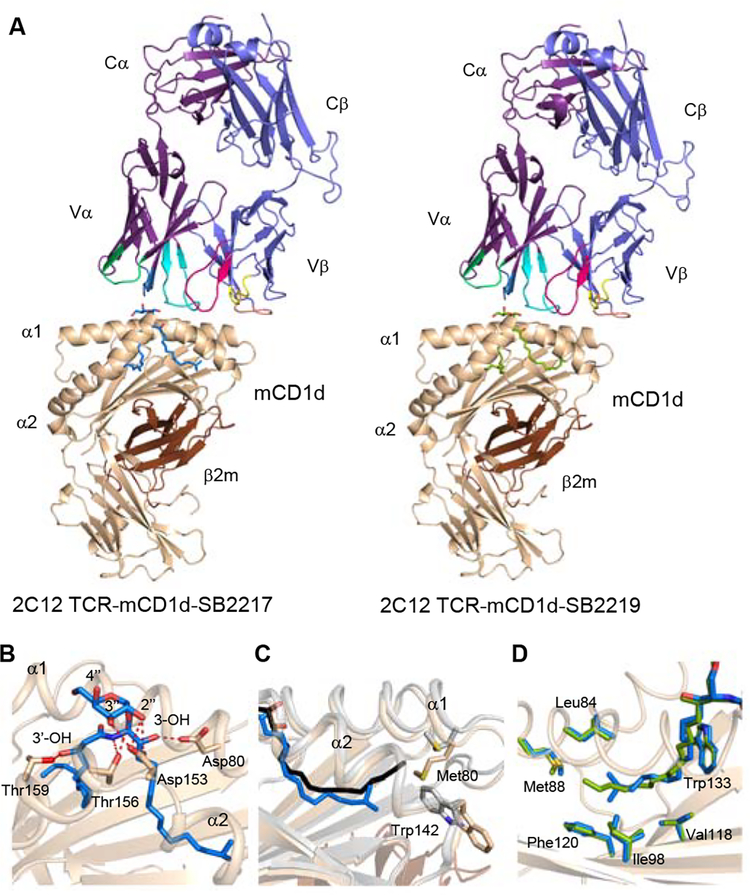
Small molecules derived from symbiotic microbiota critically contribute to intestinal immune maturation and regulation1. However, little is known about the molecular mechanisms that control immune development in the host-microbiota environment. Here, using a targeted lipidomic analysis and synthetic approach, we carried out a multifaceted investigation of immunomodulatory α-galactosylceramides from the human symbiont Bacteroides fragilis (BfaGCs). The characteristic terminal branching of BfaGCs is the result of incorporation of branched-chain amino acids taken up in the host gut by B. fragilis. A B. fragilis knockout strain that cannot metabolize branched-chain amino acids showed reduced branching in BfaGCs, and mice monocolonized with this mutant strain had impaired colonic natural killer T (NKT) cell regulation, implying structure-specific immunomodulatory activity. The sphinganine chain branching of BfaGCs is a critical determinant of NKT cell activation, which induces specific immunomodulatory gene expression signatures and effector functions. Co-crystal structure and affinity analyses of CD1d-BfaGC-NKT cell receptor complexes confirmed the interaction of BfaGCs as CD1d-restricted ligands. We present a structural and molecular-level paradigm of immunomodulatory control by interactions of endobiotic metabolites with diet, microbiota and the immune system.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
DECLARATION OF INTERESTS
S.F.O., R.S.B., and D.L.K. filed a patent on the functions of BfaGCs and related structures (US patent 10,329,315).
S.F.O., S.B.P., and D.L.K. filed a patent on the functions of BfaGCs and related structures (under review).
Figures
References
-
- Skelly AN, Sato Y, Kearney S & Honda K Mining the microbiota for microbial and metabolite-based immunotherapies. Nature Reviews Immunology 19, 305–323 (2019). - PubMed
Reference for Methods
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
