

GPCR activation mechanisms across classes and macro/microscales
- PMID: 34759375
- PMCID: PMC8580822
- DOI: 10.1038/s41594-021-00674-7
GPCR activation mechanisms across classes and macro/microscales
Abstract
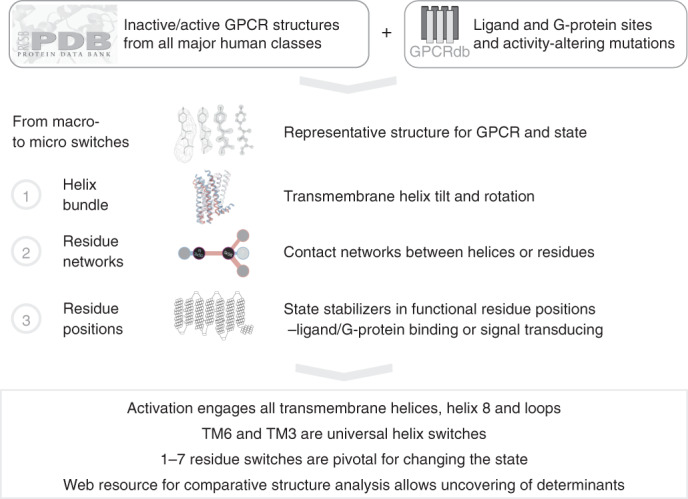
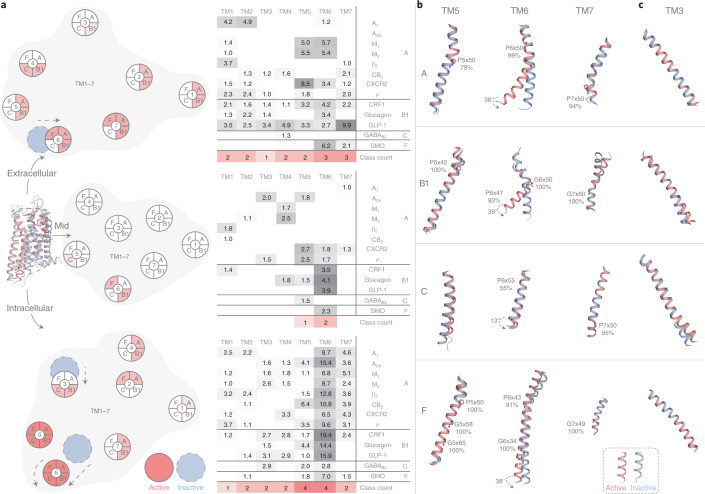
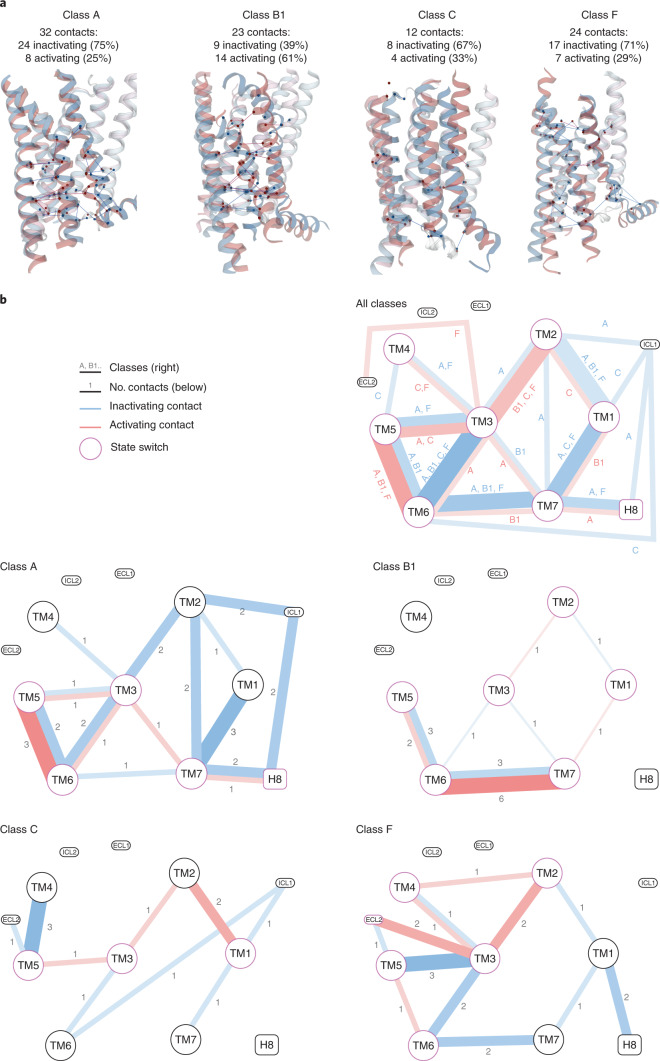
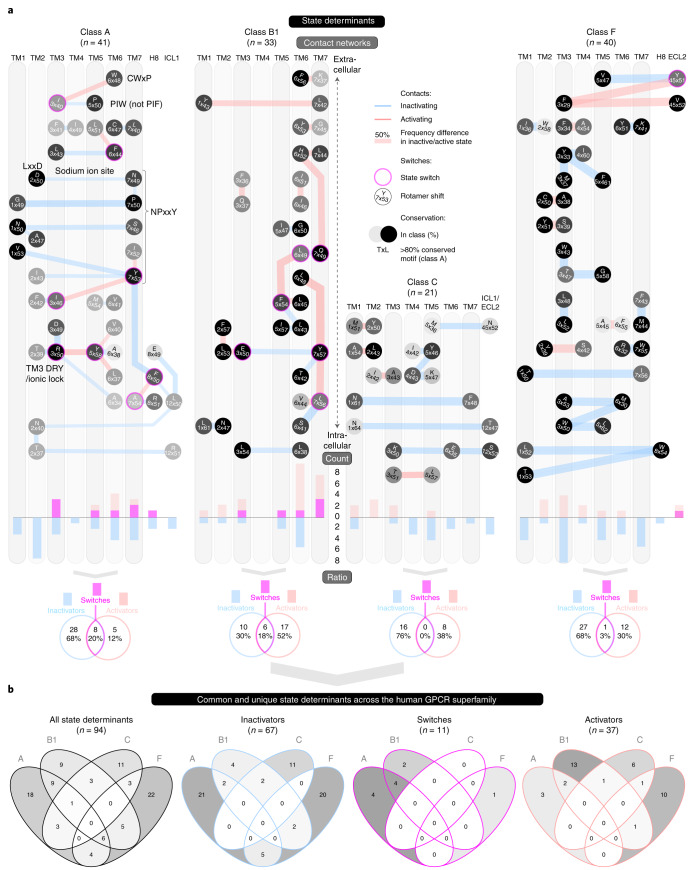
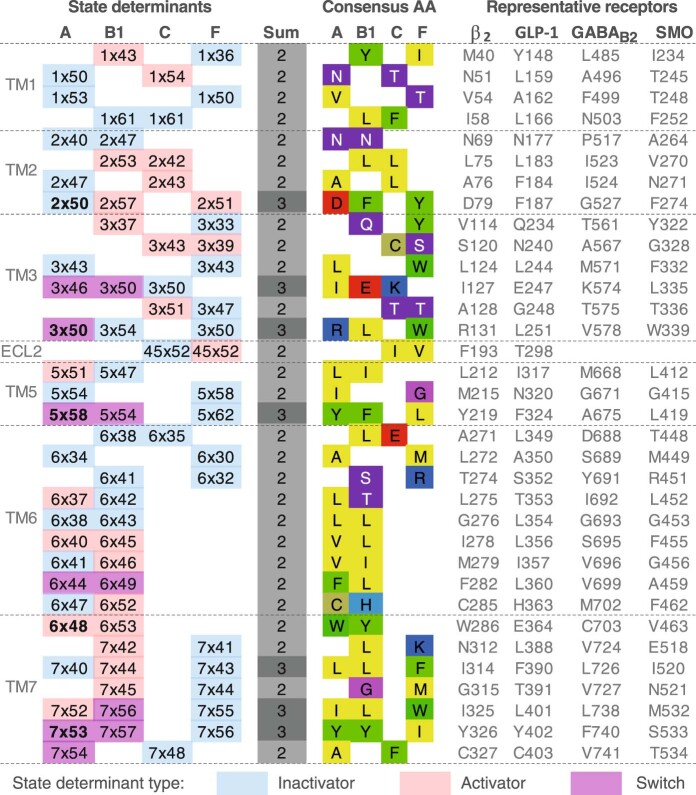
Two-thirds of human hormones and one-third of clinical drugs activate ~350 G-protein-coupled receptors (GPCR) belonging to four classes: A, B1, C and F. Whereas a model of activation has been described for class A, very little is known about the activation of the other classes, which differ by being activated by endogenous ligands bound mainly or entirely extracellularly. Here we show that, although they use the same structural scaffold and share several 'helix macroswitches', the GPCR classes differ in their 'residue microswitch' positions and contacts. We present molecular mechanistic maps of activation for each GPCR class and methods for contact analysis applicable for any functional determinants. This provides a superfamily residue-level rationale for conformational selection and allosteric communication by ligands and G proteins, laying the foundation for receptor-function studies and drugs with the desired modality.
© 2021. The Author(s).
Conflict of interest statement
M.B. is the president of Domain Therapeutics scientific advisory board. D.B.V. is a founder and a director of Z7 Biotech Ltd. After the completion of this study, C.M. moved to become an employee of Novozymes A/S. The other authors declare no competing interests.
Figures
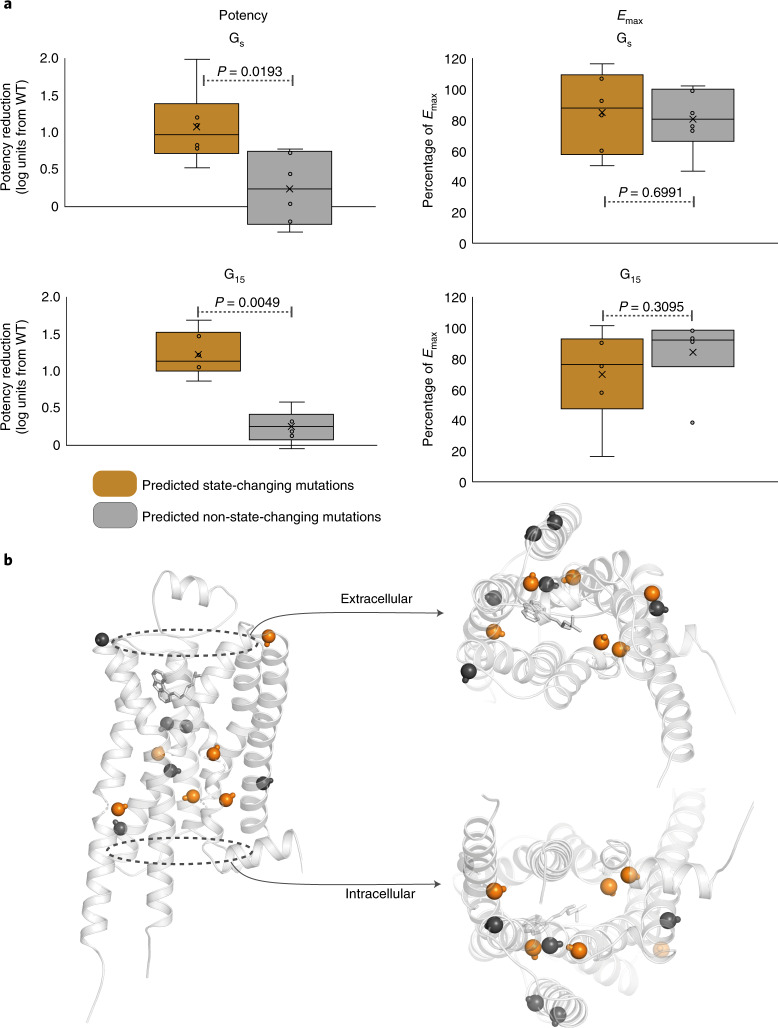
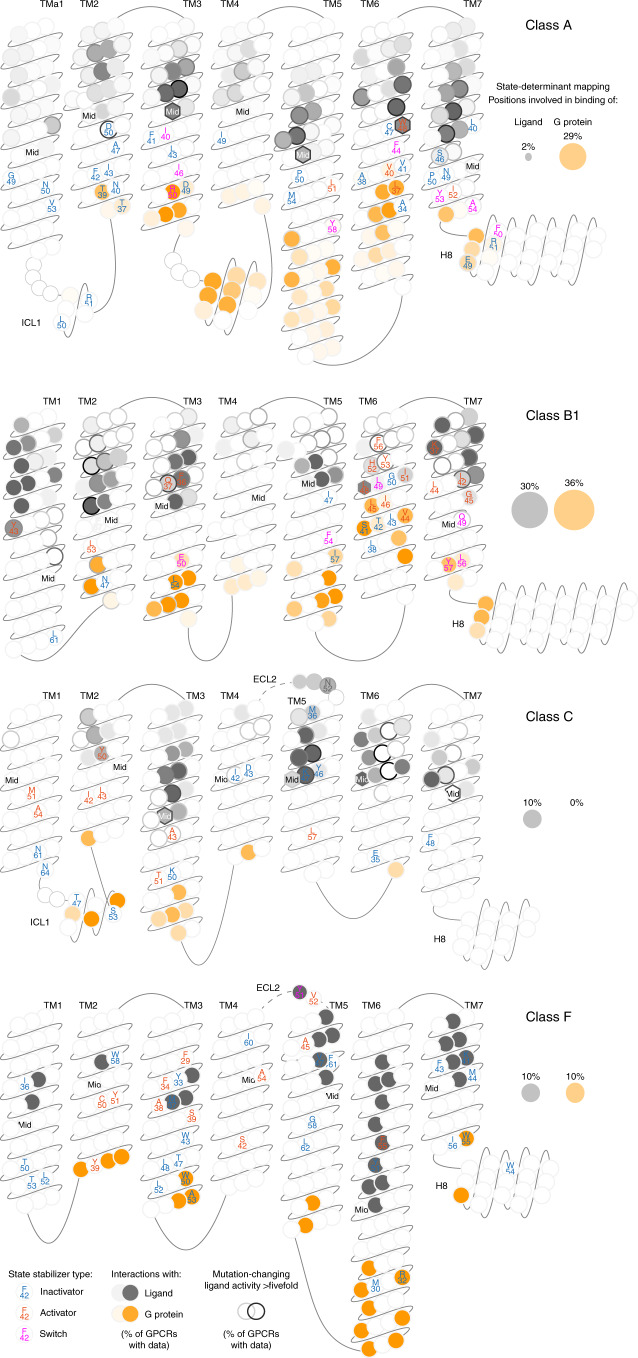
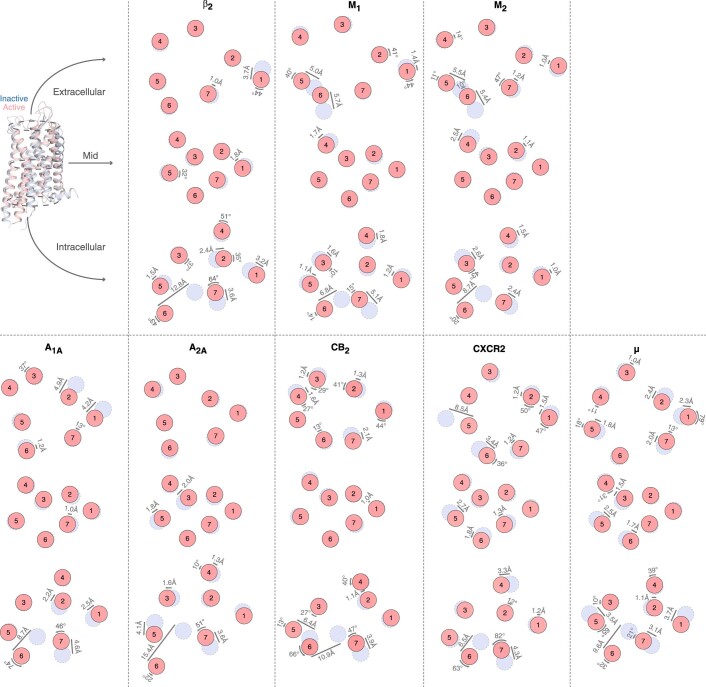
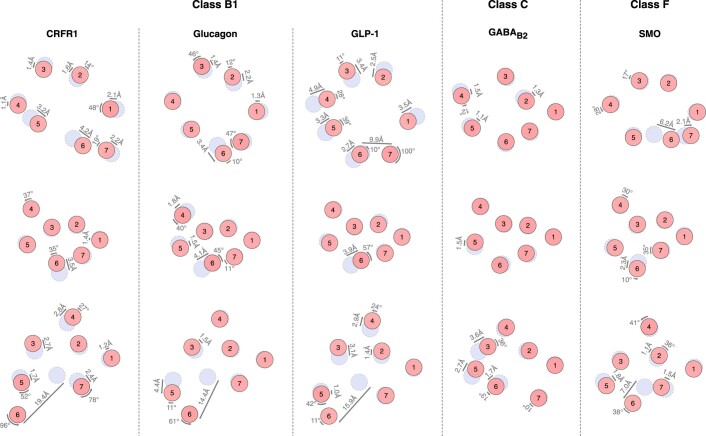
References
-
- Kolakowski LF., Jr. GCRDb: a G-protein-coupled receptor database. Recept. Channels. 1994;2:1–7. - PubMed
-
- Attwood TK, Findlay JB. Design of a discriminating fingerprint for G-protein-coupled receptors. Protein Eng. 1993;6:167–176. - PubMed
-
- Attwood TK, Findlay JB. Fingerprinting G-protein-coupled receptors. Protein Eng. 1994;7:195–203. - PubMed
-
- Fredriksson R, Lagerstrom MC, Lundin LG, Schioth HB. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003;63:1256–1272. - PubMed
-
- Nordstrom KJ, Sallman Almen M, Edstam MM, Fredriksson R, Schioth HB. Independent HHsearch, Needleman–Wunsch-based, and motif analyses reveal the overall hierarchy for most of the G protein-coupled receptor families. Mol. Biol. Evol. 2011;28:2471–2480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
