

Cellular Size, Gap Junctions, and Sodium Channel Properties Govern Developmental Changes in Cardiac Conduction
- PMID: 34759834
- PMCID: PMC8573326
- DOI: 10.3389/fphys.2021.731025
Cellular Size, Gap Junctions, and Sodium Channel Properties Govern Developmental Changes in Cardiac Conduction
Abstract
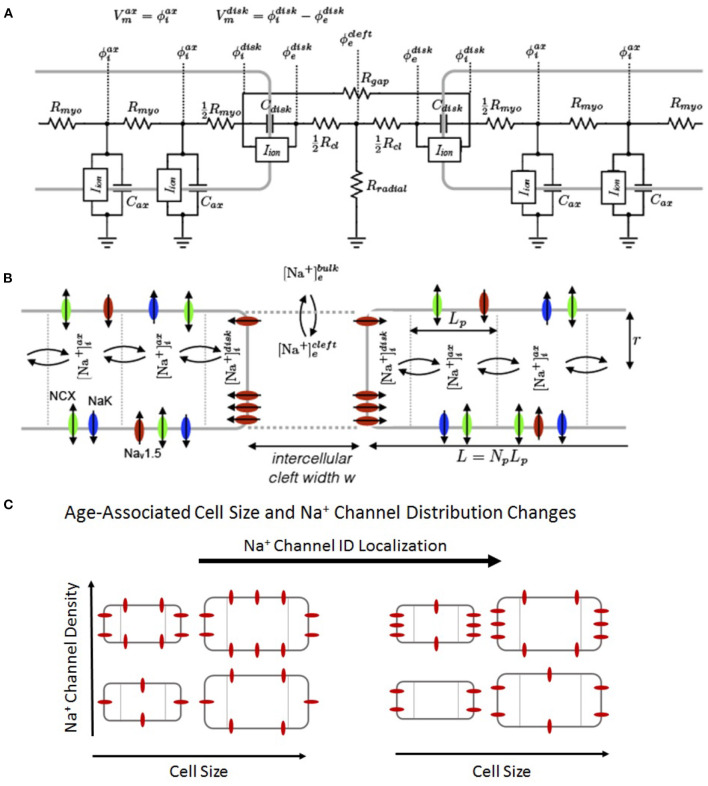
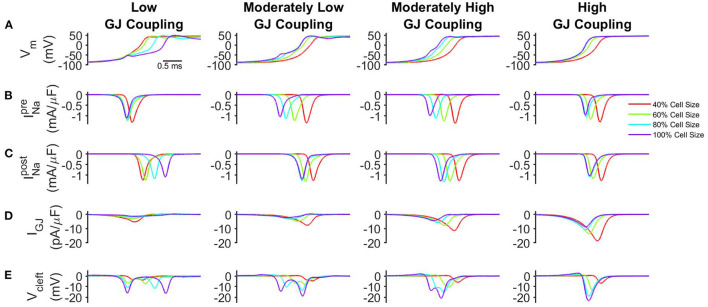
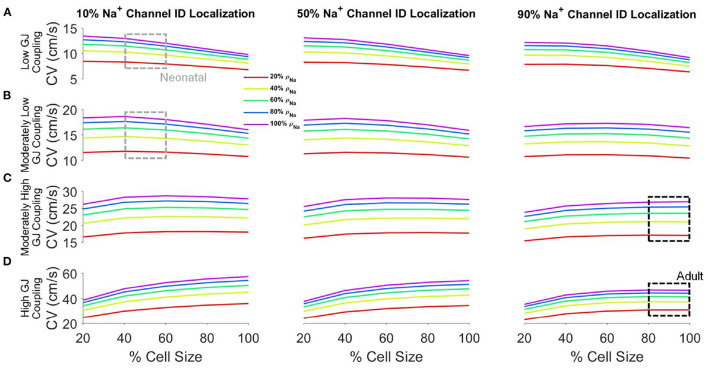
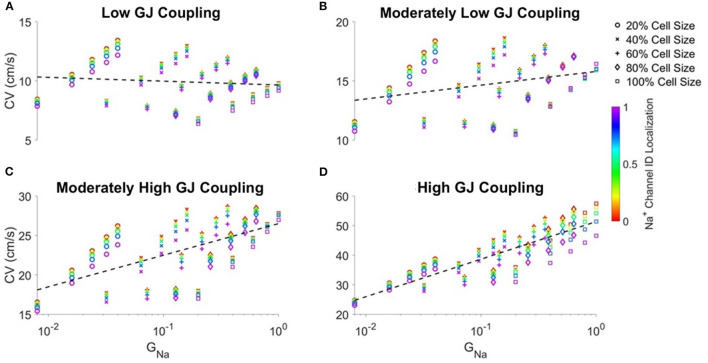
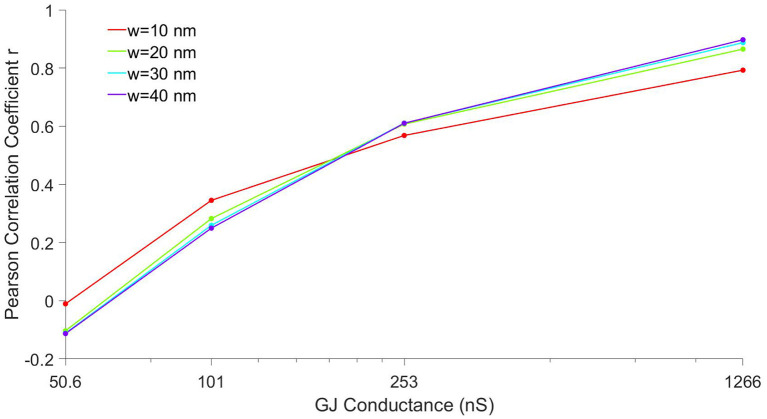
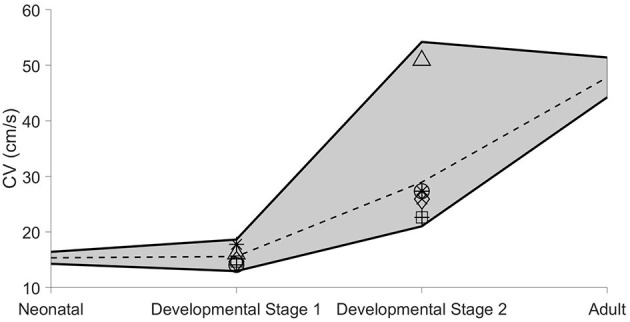
Electrical conduction in cardiac ventricular tissue is regulated via sodium (Na+) channels and gap junctions (GJs). We and others have recently shown that Na+channels preferentially localize at the site of cell-cell junctions, the intercalated disc (ID), in adult cardiac tissue, facilitating coupling via the formation of intercellular Na+nanodomains, also termed ephaptic coupling (EpC). Several properties governing EpC vary with age, including Na+channel and GJ expression and distribution and cell size. Prior work has shown that neonatal cardiomyocytes have immature IDs with Na+channels and GJs diffusively distributed throughout the sarcolemma, while adult cells have mature IDs with preferentially localized Na+channels and GJs. In this study, we perform an in silico investigation of key age-dependent properties to determine developmental regulation of cardiac conduction. Simulations predict that conduction velocity (CV) biphasically depends on cell size, depending on the strength of GJ coupling. Total cell Na+channel conductance is predictive of CV in cardiac tissue with high GJ coupling, but not correlated with CV for low GJ coupling. We find that ephaptic effects are greatest for larger cells with low GJ coupling typically associated with intermediate developmental stages. Finally, simulations illustrate how variability in cellular properties during different developmental stages can result in a range of possible CV values, with a narrow range for both neonatal and adult myocardium but a much wider range for an intermediate developmental stage. Thus, we find that developmental changes predict associated changes in cardiac conduction.
Keywords: cardiac conduction; cardiac electrophysiology; computational models; development; intercalated disc.
Copyright © 2021 Nowak, Veeraraghavan, Poelzing and Weinberg.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Agullo-Pascual E., Lin X., Leo-Macias A., Zhang M., Liang F.-X., Li Z., et al. (2014). Super-resolution imaging reveals that loss of the c-terminus of connexin43 limits microtubule plus-end capture and nav1. 5 localization at the intercalated disc. Cardiovasc. Res. 104, 371–381. 10.1093/cvr/cvu195 - DOI - PMC - PubMed
-
- Beaufort-Krol G. C., van den Berg M. P., Wilde A. A., van Tintelen J. P., Viersma J. W., Bezzina C. R., et al. (2005). Developmental aspects of long QT syndrome type 3 and Brugada syndrome on the basis of a single SCN5A mutation in childhood. J. Am. Coll. Cardiol. 46, 331–337. 10.1016/j.jacc.2005.03.066 - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous