

An exercise "sweet spot" reverses cognitive deficits of aging by growth-hormone-induced neurogenesis
- PMID: 34761193
- PMCID: PMC8567379
- DOI: 10.1016/j.isci.2021.103275
An exercise "sweet spot" reverses cognitive deficits of aging by growth-hormone-induced neurogenesis
Abstract
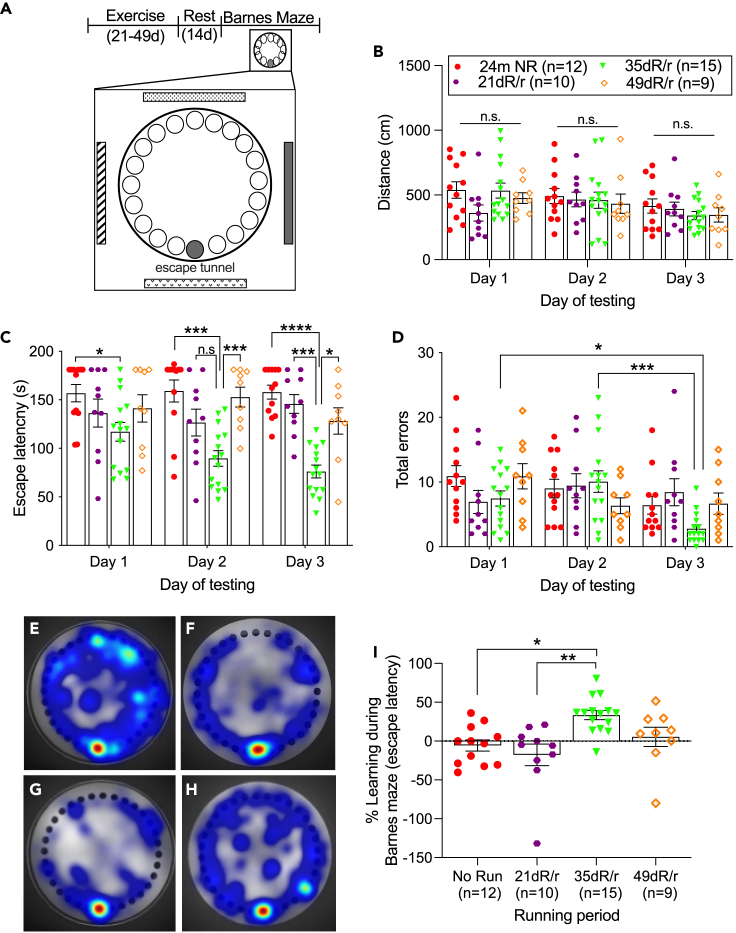
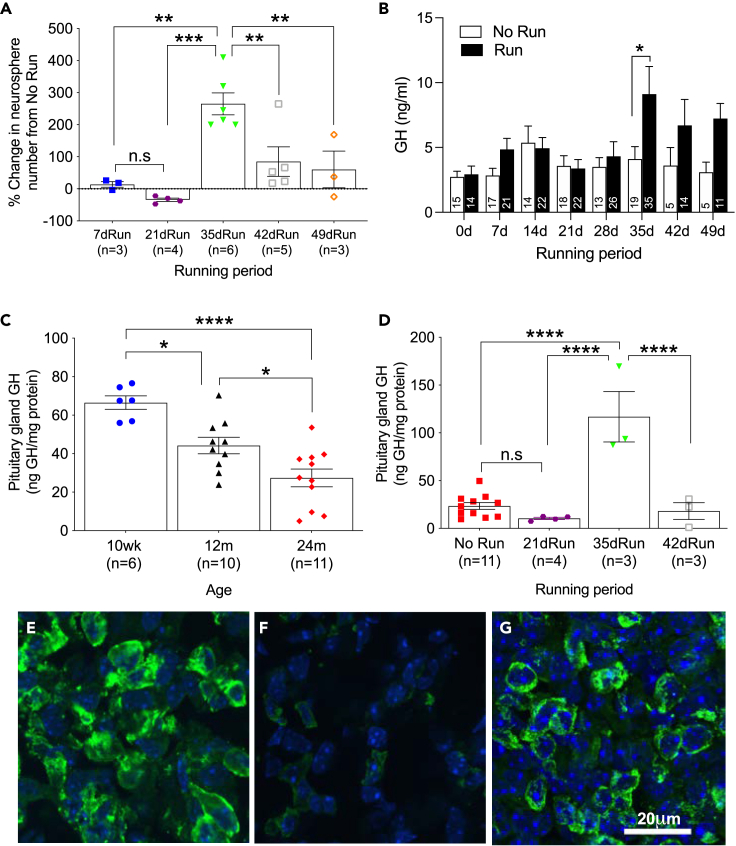
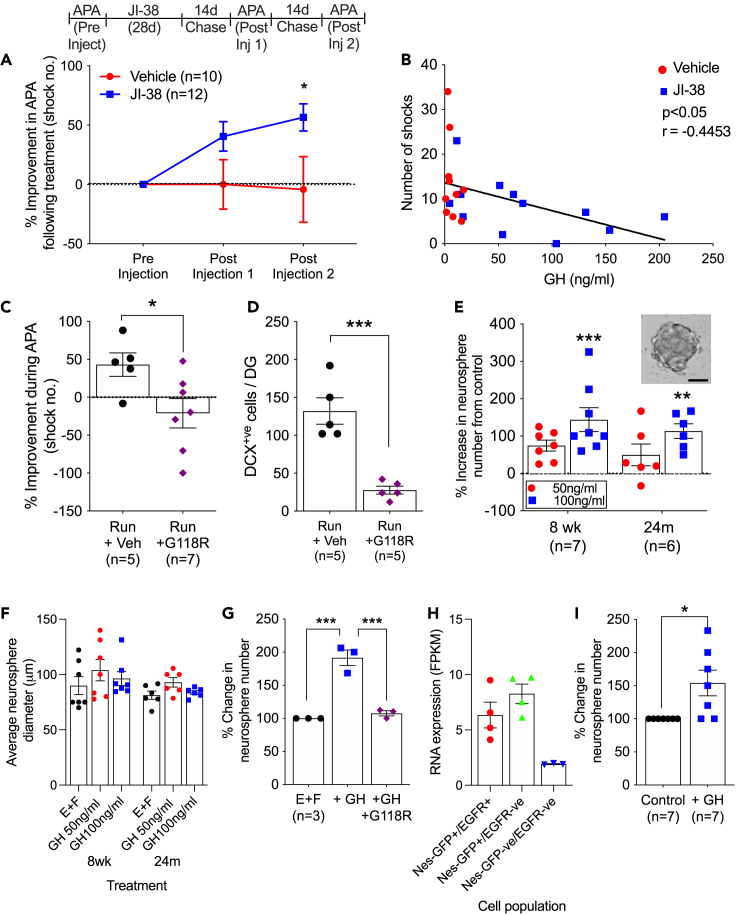
Hippocampal function is critical for spatial and contextual learning, and its decline with age contributes to cognitive impairment. Exercise can improve hippocampal function, however, the amount of exercise and mechanisms mediating improvement remain largely unknown. Here, we show exercise reverses learning deficits in aged (24 months) female mice but only when it occurs for a specific duration, with longer or shorter periods proving ineffective. A spike in the levels of growth hormone (GH) and a corresponding increase in neurogenesis during this sweet spot mediate this effect because blocking GH receptor with a competitive antagonist or depleting newborn neurons abrogates the exercise-induced cognitive improvement. Moreover, raising GH levels with GH-releasing hormone agonist improved cognition in nonrunners. We show that GH stimulates neural precursors directly, indicating the link between raised GH and neurogenesis is the basis for the substantially improved learning in aged animals.
Keywords: Age; Endocrine system physiology; Neuroscience.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Aberg D.N., Lind J., Isgaard J., Georg Kuhn H. Peripheral growth hormone induces cell proliferation in the intact adult rat brain. Growth Horm. IGF Res. 2010;20:264–269. - PubMed
-
- Ajo R., Cacicedo L., Navarro C., Sanchez-Franco F. Growth hormone action on proliferation and differentiation of cerebral cortical cells from fetal rat. Endocrinology. 2003;144:1086–1097. - PubMed
-
- Alba M., Schally A.V., Salvatori R. Partial reversibility of growth hormone (GH) deficiency in the GH-releasing hormone (GHRH) knockout mouse by postnatal treatment with a GHRH analog. Endocrinology. 2005;146:1506–1513. - PubMed
-
- Altman J., Das G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965;124:319–335. - PubMed
-
- Ben Abdallah N.M., Slomianka L., Vyssotski A.L., Lipp H.P. Early age-related changes in adult hippocampal neurogenesis in C57 mice. Neurobiol. Aging. 2010;31:151–161. - PubMed
LinkOut - more resources
Full Text Sources
