

ATRX regulates glial identity and the tumor microenvironment in IDH-mutant glioma
- PMID: 34763709
- PMCID: PMC8588616
- DOI: 10.1186/s13059-021-02535-4
ATRX regulates glial identity and the tumor microenvironment in IDH-mutant glioma
Abstract
Background: Recent single-cell transcriptomic studies report that IDH-mutant gliomas share a common hierarchy of cellular phenotypes, independent of genetic subtype. However, the genetic differences between IDH-mutant glioma subtypes are prognostic, predictive of response to chemotherapy, and correlate with distinct tumor microenvironments.
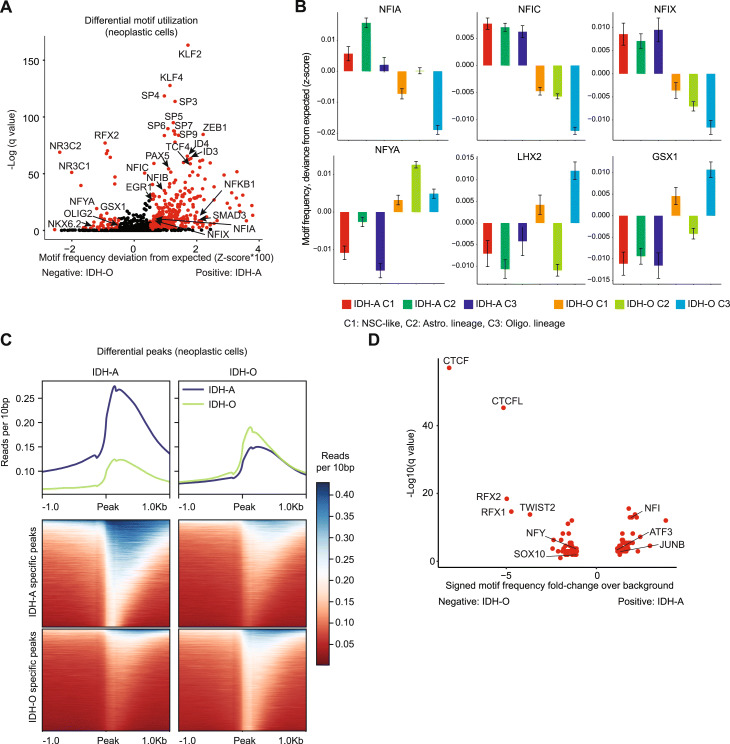
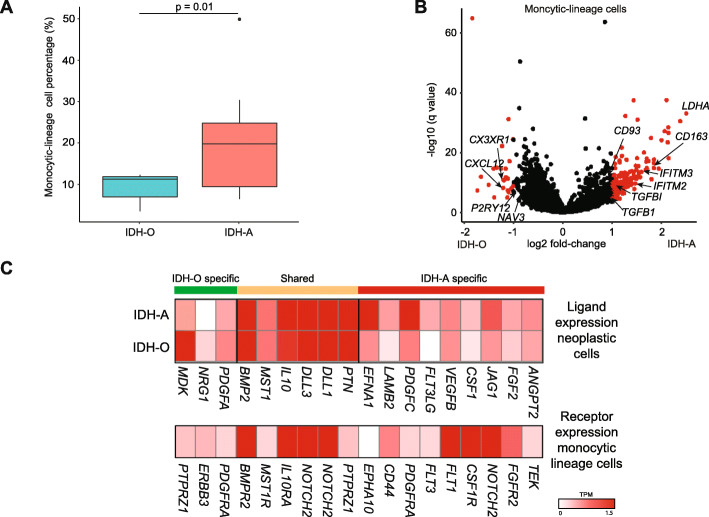
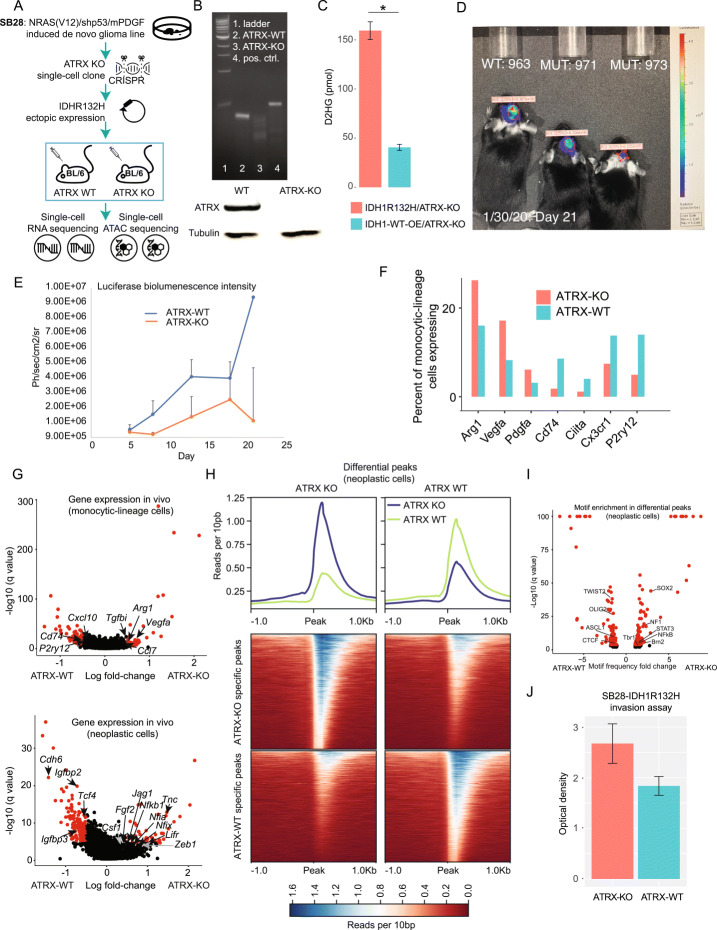
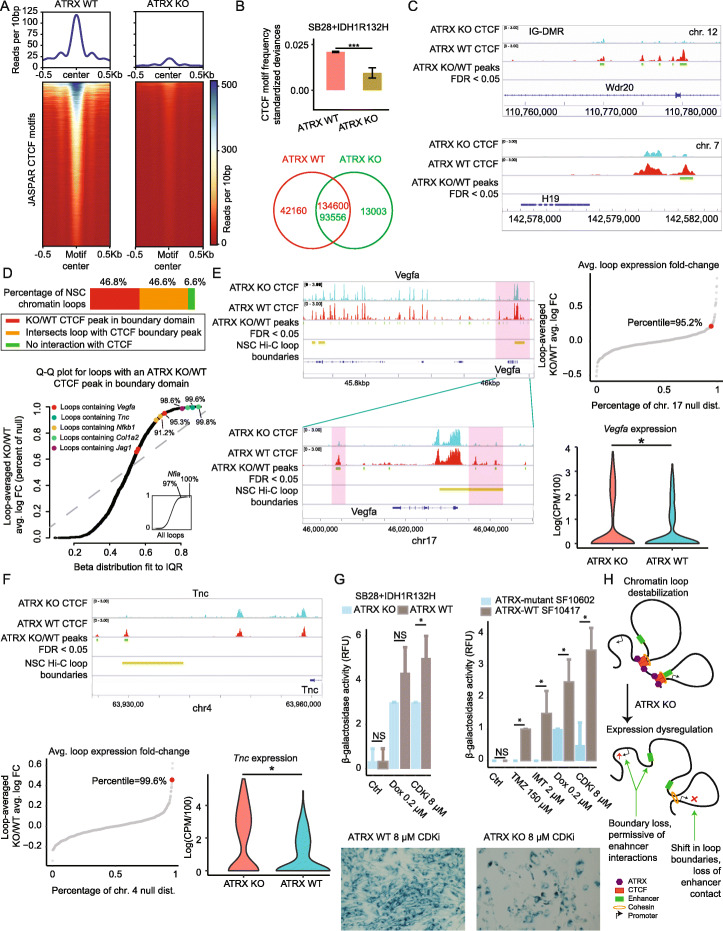
Results: To reconcile these findings, we profile 22 human IDH-mutant gliomas using scATAC-seq and scRNA-seq. We determine the cell-type-specific differences in transcription factor expression and associated regulatory grammars between IDH-mutant glioma subtypes. We find that while IDH-mutant gliomas do share a common distribution of cell types, there are significant differences in the expression and targeting of transcription factors that regulate glial identity and cytokine elaboration. We knock out the chromatin remodeler ATRX, which suffers loss-of-function alterations in most IDH-mutant astrocytomas, in an IDH-mutant immunocompetent intracranial murine model. We find that both human ATRX-mutant gliomas and murine ATRX-knockout gliomas are more heavily infiltrated by immunosuppressive monocytic-lineage cells derived from circulation than ATRX-intact gliomas, in an IDH-mutant background. ATRX knockout in murine glioma recapitulates gene expression and open chromatin signatures that are specific to human ATRX-mutant astrocytomas, including drivers of astrocytic lineage and immune-cell chemotaxis. Through single-cell cleavage under targets and tagmentation assays and meta-analysis of public data, we show that ATRX loss leads to a global depletion in CCCTC-binding factor association with DNA, gene dysregulation along associated chromatin loops, and protection from therapy-induced senescence.
Conclusions: These studies explain how IDH-mutant gliomas from different subtypes maintain distinct phenotypes and tumor microenvironments despite a common lineage hierarchy.
© 2021. The Author(s).
Conflict of interest statement
None declared.
Figures

Similar articles
-
ATRX immunostaining predicts IDH and H3F3A status in gliomas.Acta Neuropathol Commun. 2016 Jun 16;4(1):60. doi: 10.1186/s40478-016-0331-6. Acta Neuropathol Commun. 2016. PMID: 27311324 Free PMC article.
-
Immunohistochemical ATRX expression is not a surrogate for 1p19q codeletion.Brain Tumor Pathol. 2018 Apr;35(2):106-113. doi: 10.1007/s10014-018-0312-5. Epub 2018 Mar 17. Brain Tumor Pathol. 2018. PMID: 29550880
-
DNA methylation signatures for 2016 WHO classification subtypes of diffuse gliomas.Clin Epigenetics. 2017 Apr 4;9:32. doi: 10.1186/s13148-017-0331-9. eCollection 2017. Clin Epigenetics. 2017. PMID: 28392842 Free PMC article.
-
Correlation between IDH, ATRX, and TERT promoter mutations in glioma.Brain Tumor Pathol. 2020 Apr;37(2):33-40. doi: 10.1007/s10014-020-00360-4. Epub 2020 Mar 29. Brain Tumor Pathol. 2020. PMID: 32227259 Review.
-
Integrated diagnostics of diffuse astrocytic and oligodendroglial tumors.Pathologe. 2019 Jun;40(Suppl 1):9-17. doi: 10.1007/s00292-019-0581-8. Pathologe. 2019. PMID: 31025086 Review. English.
Cited by
-
Stalled oligodendrocyte differentiation in IDH-mutant gliomas.Genome Med. 2023 Apr 13;15(1):24. doi: 10.1186/s13073-023-01175-6. Genome Med. 2023. PMID: 37055795 Free PMC article.
-
Inhibitory Effect of miR-339-5p on Glioma through PTP4A1/HMGB1 Pathway.Dis Markers. 2022 Jul 15;2022:2231195. doi: 10.1155/2022/2231195. eCollection 2022. Dis Markers. 2022. PMID: 35872698 Free PMC article.
-
An immune signature to predict the prognosis of ATRX-wildtype glioma patients and guide immune checkpoint blockade therapy.Aging (Albany NY). 2023 Oct 6;15(19):10453-10472. doi: 10.18632/aging.205088. Epub 2023 Oct 6. Aging (Albany NY). 2023. PMID: 37812190 Free PMC article.
-
Deciphering the generation of heterogeneity in esophageal squamous cell carcinoma metastasis via single-cell multiomics analysis.J Transl Med. 2025 Feb 4;23(1):148. doi: 10.1186/s12967-025-06154-6. J Transl Med. 2025. PMID: 39905485 Free PMC article.
-
New insights into the Immune TME of adult-type diffuse gliomas.Curr Opin Neurol. 2022 Dec 1;35(6):794-802. doi: 10.1097/WCO.0000000000001112. Epub 2022 Oct 7. Curr Opin Neurol. 2022. PMID: 36226710 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
