

Leveraging machine learning essentiality predictions and chemogenomic interactions to identify antifungal targets
- PMID: 34764269
- PMCID: PMC8586148
- DOI: 10.1038/s41467-021-26850-3
Leveraging machine learning essentiality predictions and chemogenomic interactions to identify antifungal targets
Abstract
Fungal pathogens pose a global threat to human health, with Candida albicans among the leading killers. Systematic analysis of essential genes provides a powerful strategy to discover potential antifungal targets. Here, we build a machine learning model to generate genome-wide gene essentiality predictions for C. albicans and expand the largest functional genomics resource in this pathogen (the GRACE collection) by 866 genes. Using this model and chemogenomic analyses, we define the function of three uncharacterized essential genes with roles in kinetochore function, mitochondrial integrity, and translation, and identify the glutaminyl-tRNA synthetase Gln4 as the target of N-pyrimidinyl-β-thiophenylacrylamide (NP-BTA), an antifungal compound.
© 2021. The Author(s).
Conflict of interest statement
L.E.C. is a co-founder and shareholder in Bright Angel Therapeutics, a platform company for the development of novel antifungal therapeutics. L.E.C. is a consultant for Boragen, a small-molecule development company focused on leveraging the unique chemical properties of boron chemistry for crop protection and animal health. All other authors declare no competing interests.
Figures



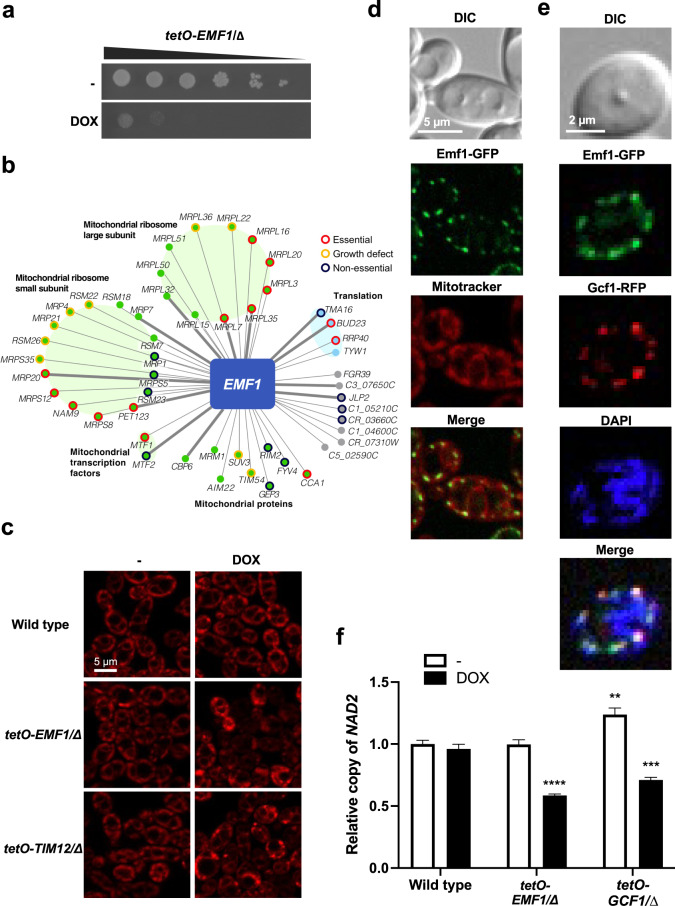


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
