

Diverse human astrocyte and microglial transcriptional responses to Alzheimer's pathology
- PMID: 34767070
- PMCID: PMC8732962
- DOI: 10.1007/s00401-021-02372-6
Diverse human astrocyte and microglial transcriptional responses to Alzheimer's pathology
Abstract
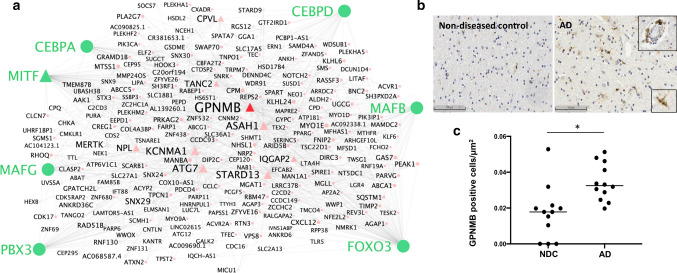
To better define roles that astrocytes and microglia play in Alzheimer's disease (AD), we used single-nuclei RNA-sequencing to comprehensively characterise transcriptomes in astrocyte and microglia nuclei selectively enriched during isolation post-mortem from neuropathologically defined AD and control brains with a range of amyloid-beta and phospho-tau (pTau) pathology. Significant differences in glial gene expression (including AD risk genes expressed in both the astrocytes [CLU, MEF2C, IQCK] and microglia [APOE, MS4A6A, PILRA]) were correlated with tissue amyloid or pTau expression. The differentially expressed genes were distinct between with the two cell types and pathologies, although common (but cell-type specific) gene sets were enriched with both pathologies in each cell type. Astrocytes showed enrichment for proteostatic, inflammatory and metal ion homeostasis pathways. Pathways for phagocytosis, inflammation and proteostasis were enriched in microglia and perivascular macrophages with greater tissue amyloid, but IL1-related pathway enrichment was found specifically in association with pTau. We also found distinguishable sub-clusters in the astrocytes and microglia characterised by transcriptional signatures related to either homeostatic functions or disease pathology. Gene co-expression analyses revealed potential functional associations of soluble biomarkers of AD in astrocytes (CLU) and microglia (GPNMB). Our work highlights responses of both astrocytes and microglia for pathological protein clearance and inflammation, as well as glial transcriptional diversity in AD.
Keywords: Alzheimer’s disease; Amyloid-beta; Astrocytes; Microglia; Tau; snRNA sequencing.
© 2021. The Author(s).
Conflict of interest statement
PMM has received consultancy fees from Roche, Adelphi Communications, Celgene, Neurodiem, and Medscape. He has received honoraria or speakers’ fees from Novartis and Biogen and has received research or educational funds from Biogen, Novartis, and GlaxoSmithKline.
Figures




References
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc: Ser B (Methodol) 1995;57:289–300.
Publication types
MeSH terms
Grants and funding
- MR/N026934/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_17114/MRC_/Medical Research Council/United Kingdom
- MR/M024903/1/MRC_/Medical Research Council/United Kingdom
- G0601966/MRC_/Medical Research Council/United Kingdom
- MR/N008219/1/MRC_/Medical Research Council/United Kingdom
- G0100811/MRC_/Medical Research Council/United Kingdom
- MR/K501013/1/MRC_/Medical Research Council/United Kingdom
- DH_/Department of Health/United Kingdom
- G0300665/MRC_/Medical Research Council/United Kingdom
- MR/T031891/1/MRC_/Medical Research Council/United Kingdom
- G9901399/MRC_/Medical Research Council/United Kingdom
- G9409531/MRC_/Medical Research Council/United Kingdom
- G0900897/MRC_/Medical Research Council/United Kingdom
- G9409634/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
