

Lactate supports a metabolic-epigenetic link in macrophage polarization
- PMID: 34767443
- PMCID: PMC8589316
- DOI: 10.1126/sciadv.abi8602
Lactate supports a metabolic-epigenetic link in macrophage polarization
Abstract
Lactate accumulation is a hallmark of solid cancers and is linked to the immune suppressive phenotypes of tumor-infiltrating immune cells. We report herein that interleukin-4 (IL-4)–induced M0 → M2 macrophage polarization is accompanied by interchangeable glucose- or lactate-dependent tricarboxylic acid (TCA) cycle metabolism that directly drives histone acetylation, M2 gene transcription, and functional immune suppression. Lactate-dependent M0 → M2 polarization requires both mitochondrial pyruvate uptake and adenosine triphosphate–citrate lyase (ACLY) enzymatic activity. Notably, exogenous acetate rescues defective M2 polarization and histone acetylation following mitochondrial pyruvate carrier 1 (MPC1) inhibition or ACLY deficiency. Lastly, M2 macrophage–dependent tumor progression is impaired by conditional macrophage ACLY deficiency, further supporting a dominant role for glucose/lactate mitochondrial metabolism and histone acetylation in driving immune evasion. This work adds to our understanding of how mitochondrial metabolism affects macrophage functional phenotypes and identifies a unique tumor microenvironment (TME)–driven metabolic-epigenetic link in M2 macrophages.
Figures
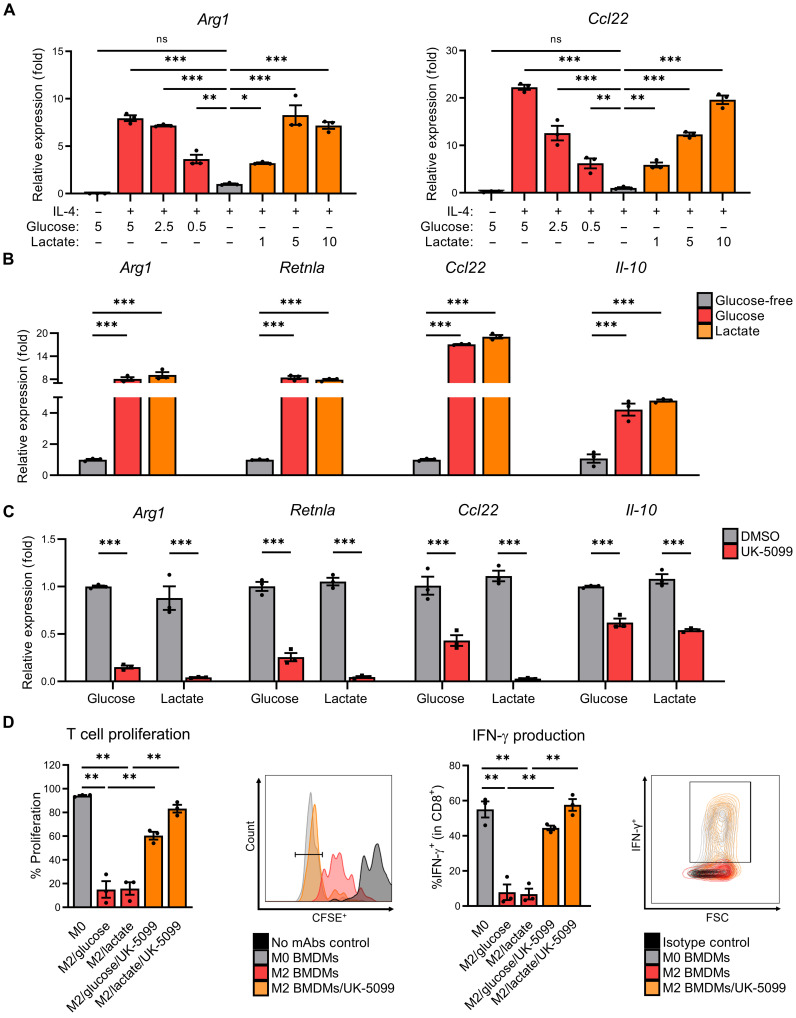
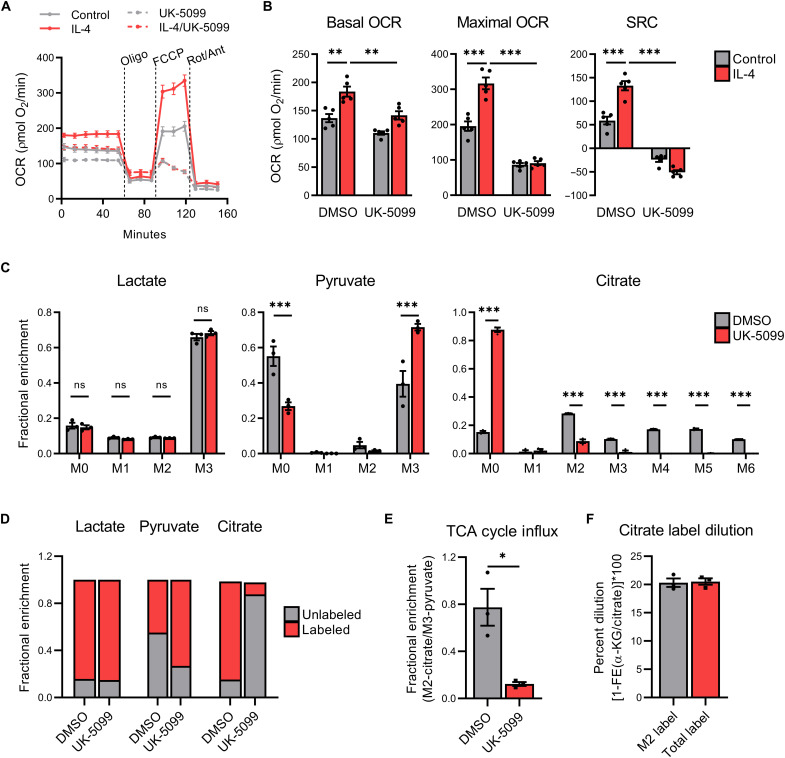
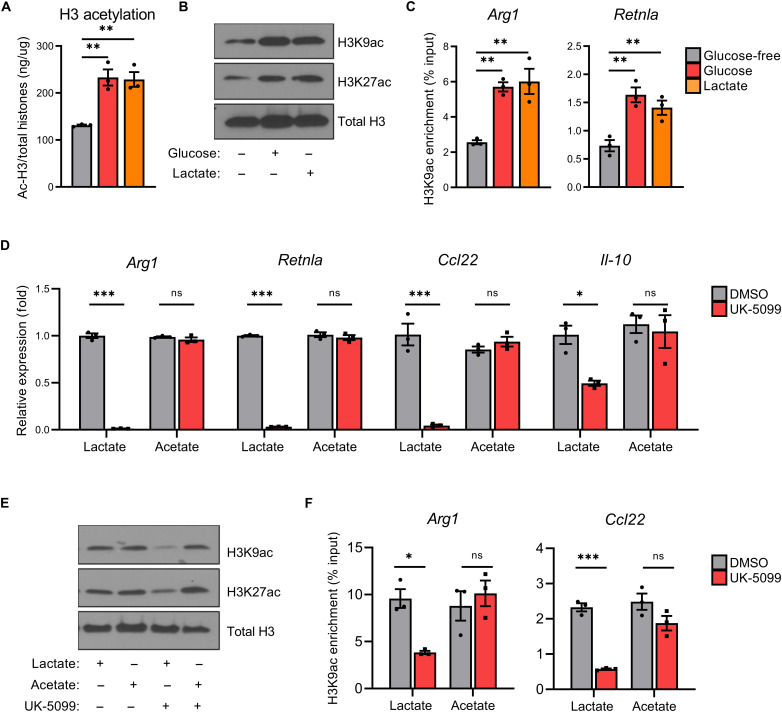
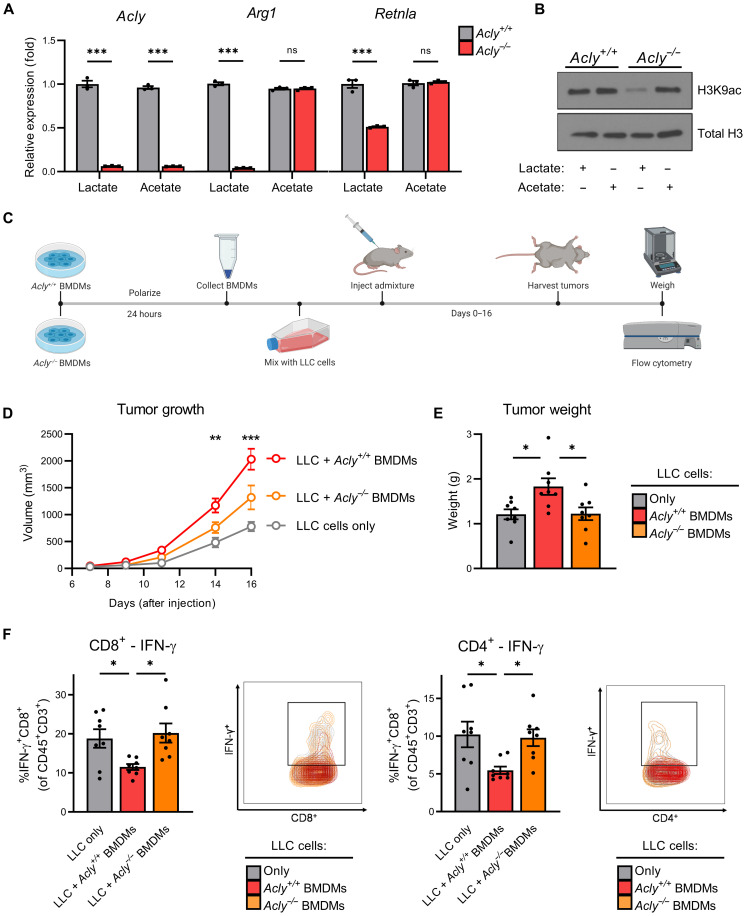
References
-
- Warburg O., On the origin of cancer cells. Science 123, 309–314 (1956). - PubMed
-
- Warburg O., On respiratory impairment in cancer cells. Science 124, 269–270 (1956). - PubMed
-
- Certo M., Tsai C.-H., Pucino V., Ho P.-C., Mauro C., Lactate modulation of immune responses in inflammatory versus tumour microenvironments. Nat. Rev. Immunol. 21, 151–161 (2021). - PubMed
-
- Angelin A., Gil-de-Gómez L., Dahiya S., Jiao J., Guo L., Levine M. H., Wang Z., Quinn W. J. III, Kopinski P. K., Wang L., Akimova T., Liu Y., Bhatti T. R., Han R., Laskin B. L., Baur J. A., Blair I. A., Wallace D. C., Hancock W. W., Beier U. H., Foxp3 reprograms T cell metabolism to function in low-glucose, high-lactate environments. Cell Metab. 25, 1282–1293.e7 (2017). - PMC - PubMed
-
- Zhang D., Tang Z., Huang H., Zhou G., Cui C., Weng Y., Liu W., Kim S., Lee S., Perez-Neut M., Ding J., Czyz D., Hu R., Ye Z., He M., Zheng Y. G., Shuman H. A., Dai L., Ren B., Roeder R. G., Becker L., Zhao Y., Metabolic regulation of gene expression by histone lactylation. Nature 574, 575–580 (2019). - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
