

Single-cell multiomics defines tolerogenic extrathymic Aire-expressing populations with unique homology to thymic epithelium
- PMID: 34767455
- PMCID: PMC8855935
- DOI: 10.1126/sciimmunol.abl5053
Single-cell multiomics defines tolerogenic extrathymic Aire-expressing populations with unique homology to thymic epithelium
Abstract
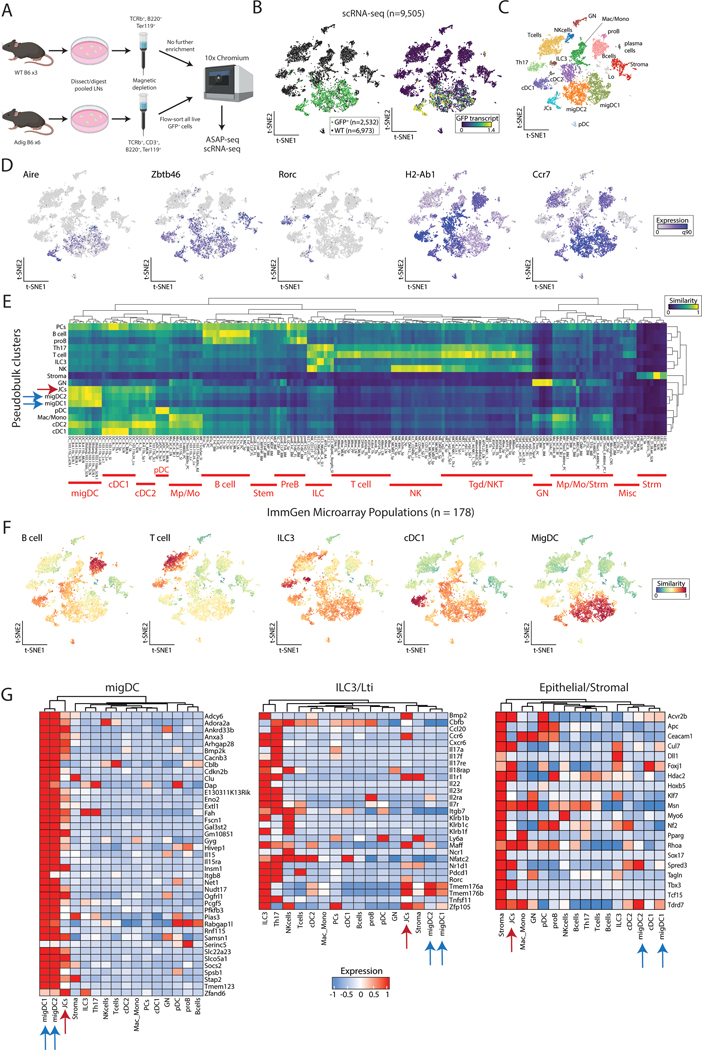
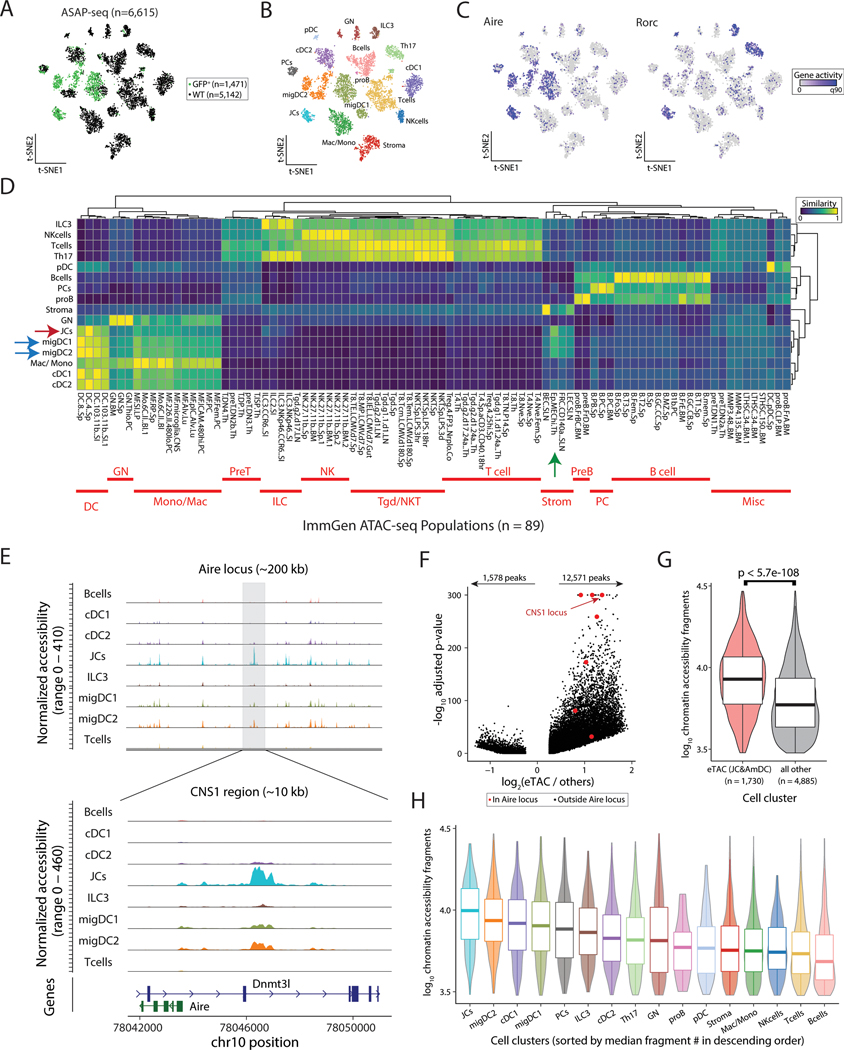
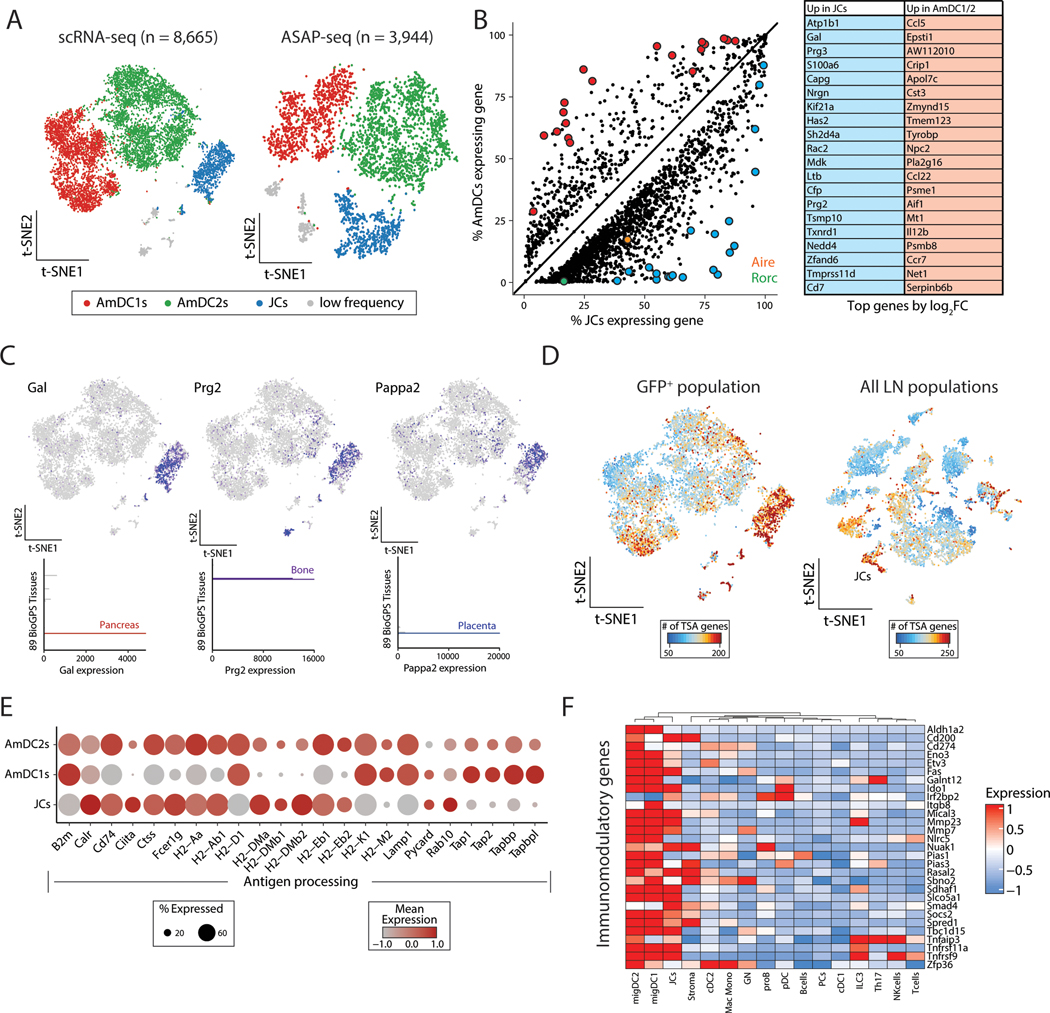
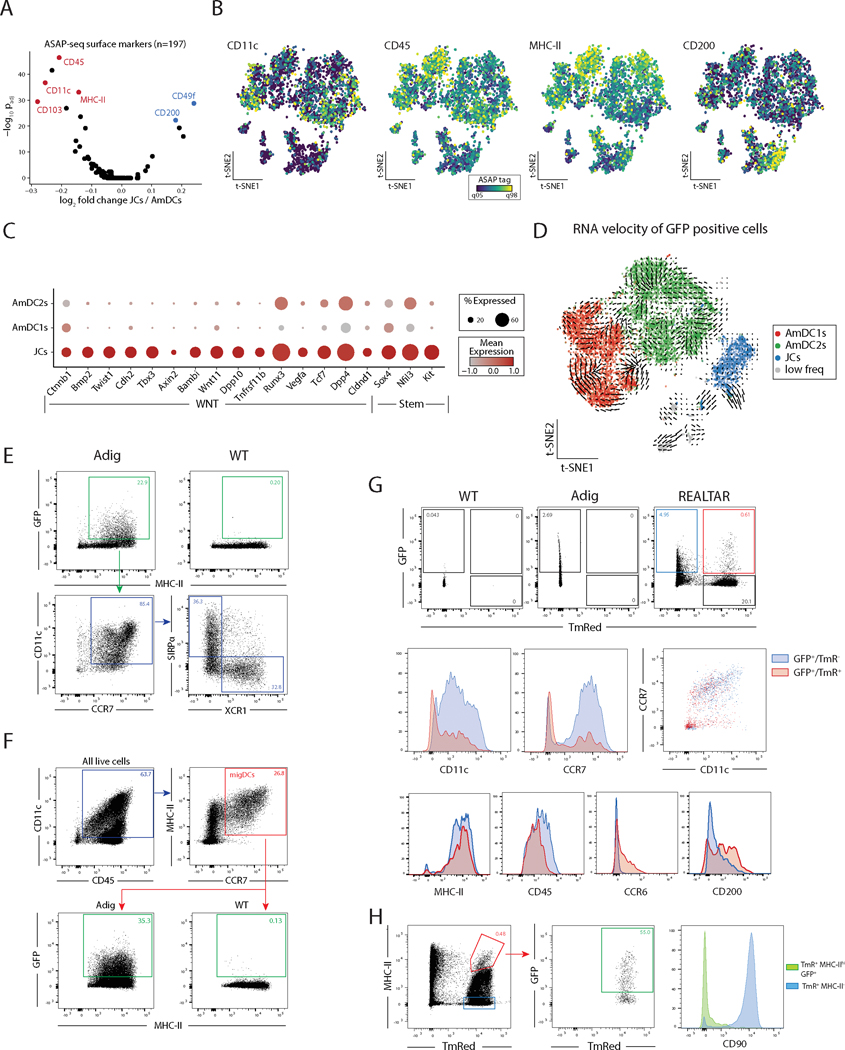
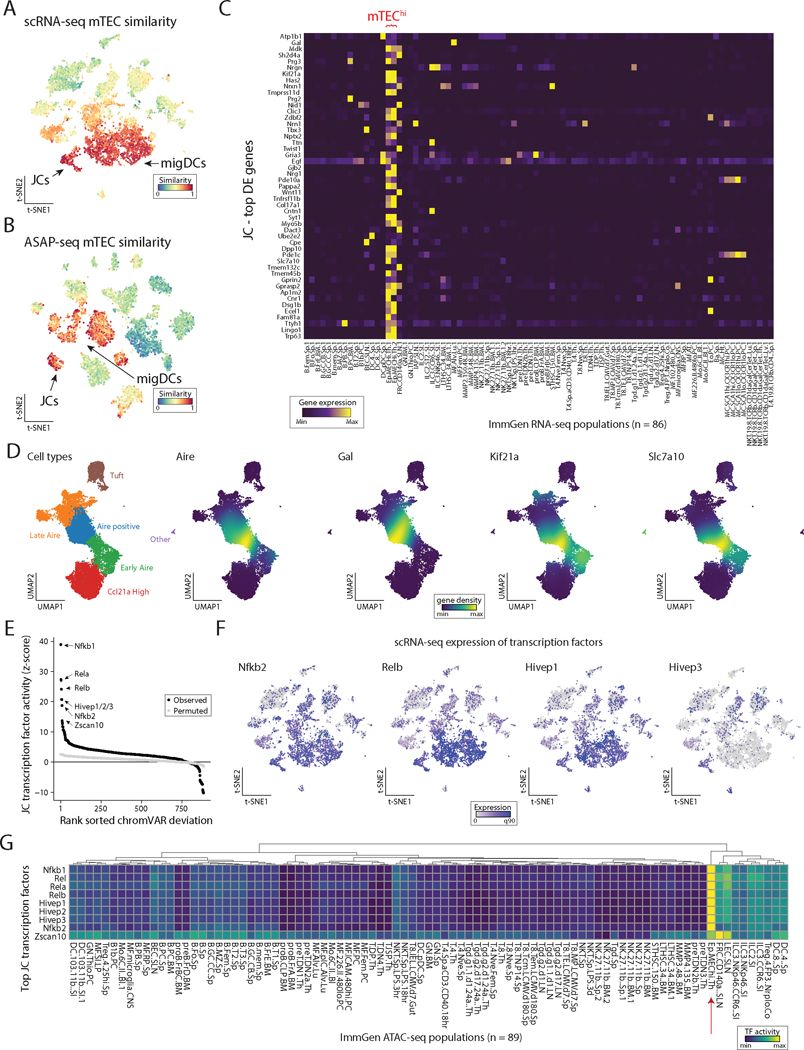
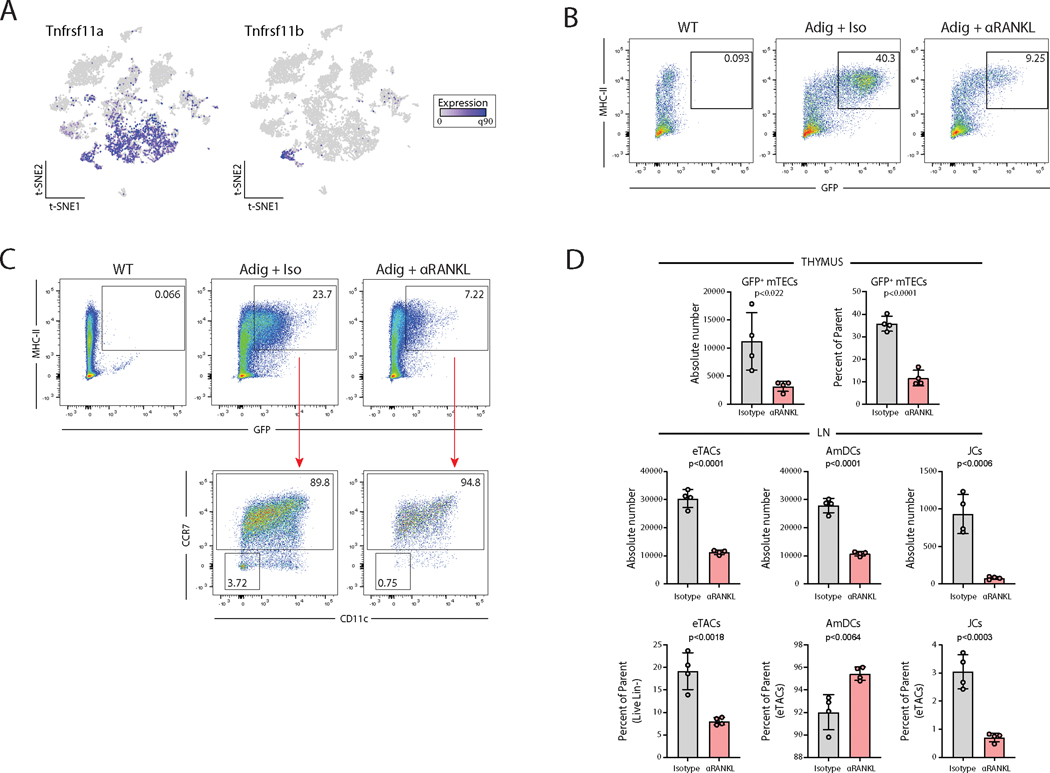
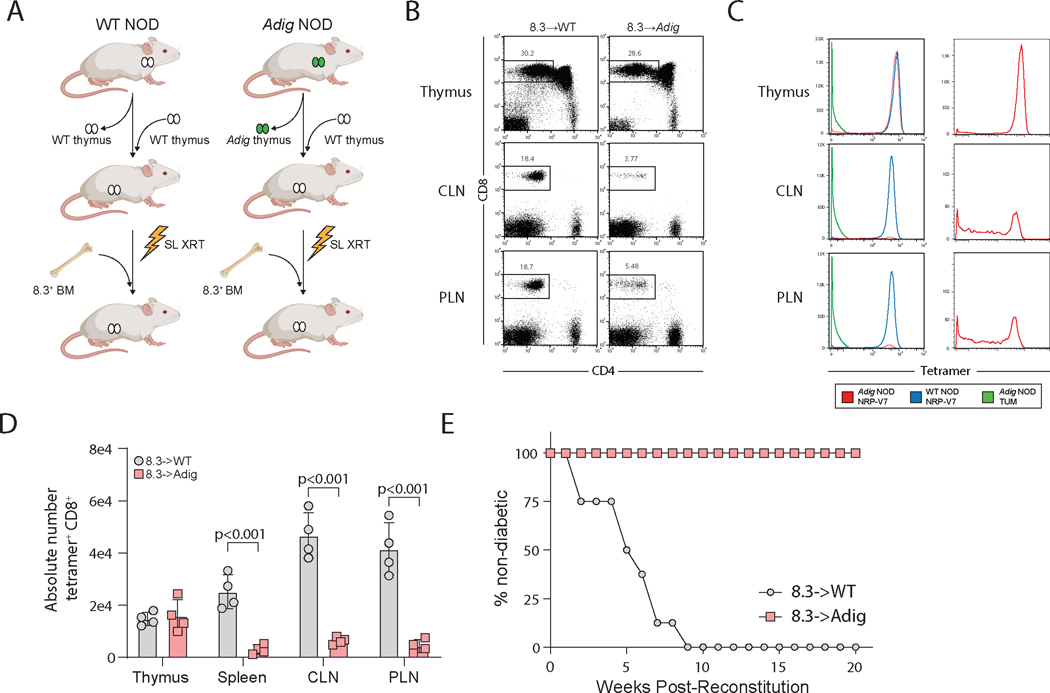
The autoimmune regulator (Aire), a well-defined transcriptional regulator in the thymus, is also found in extrathymic Aire-expressing cells (eTACs) in the secondary lymphoid organs. eTACs are hematopoietic antigen-presenting cells and inducers of immune tolerance, but their precise identity has remained unclear. Here, we use single-cell multiomics, transgenic murine models, and functional approaches to define eTACs at the transcriptional, genomic, and proteomic level. We find that eTACs consist of two similar cell types: CCR7+ Aire-expressing migratory dendritic cells (AmDCs) and an Airehi population coexpressing Aire and retinoic acid receptor–related orphan receptor γt (RORγt) that we term Janus cells (JCs). Both JCs and AmDCs have the highest transcriptional and genomic homology to CCR7+ migratory dendritic cells. eTACs, particularly JCs, have highly accessible chromatin and broad gene expression, including a range of tissue-specific antigens, as well as remarkable homology to medullary thymic epithelium and RANK-dependent Aire expression. Transgenic self-antigen expression by eTACs is sufficient to induce negative selection and prevent autoimmune diabetes. This transcriptional, genomic, and functional symmetry between eTACs (both JCs and AmDCs) and medullary thymic epithelium—the other principal Aire-expressing population and a key regulator of central tolerance—identifies a core program that may influence self-representation and tolerance across the spectrum of immune development.
Figures
References
-
- Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, et al. Projection of an immunological self shadow within the thymus by the aire protein. Science (80- ). 2002; - PubMed
-
- Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D. The cellular mechanism of Aire control of T cell tolerance. Immunity. 2005;23(2):227–39. - PubMed
-
- Ramsey C, Hässler S, Marits P, Kämpe O, Surh CD, Peltonen L, et al. Increased antigen presenting cell-mediated T cell activation in mice and patients without the autoimmune regulator. Eur J Immunol. 2006;36(2):305–17. - PubMed
