

FERONIA and microtubules independently contribute to mechanical integrity in the Arabidopsis shoot
- PMID: 34767544
- PMCID: PMC8612563
- DOI: 10.1371/journal.pbio.3001454
FERONIA and microtubules independently contribute to mechanical integrity in the Arabidopsis shoot
Abstract
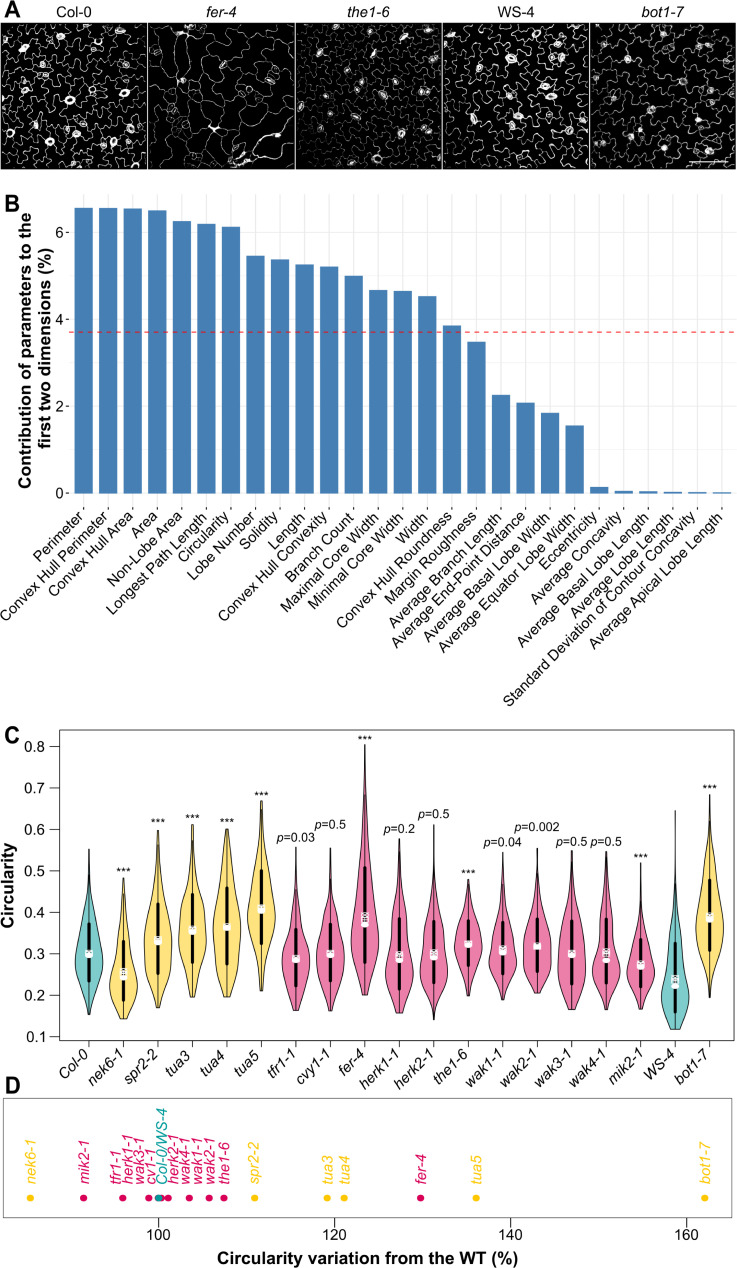
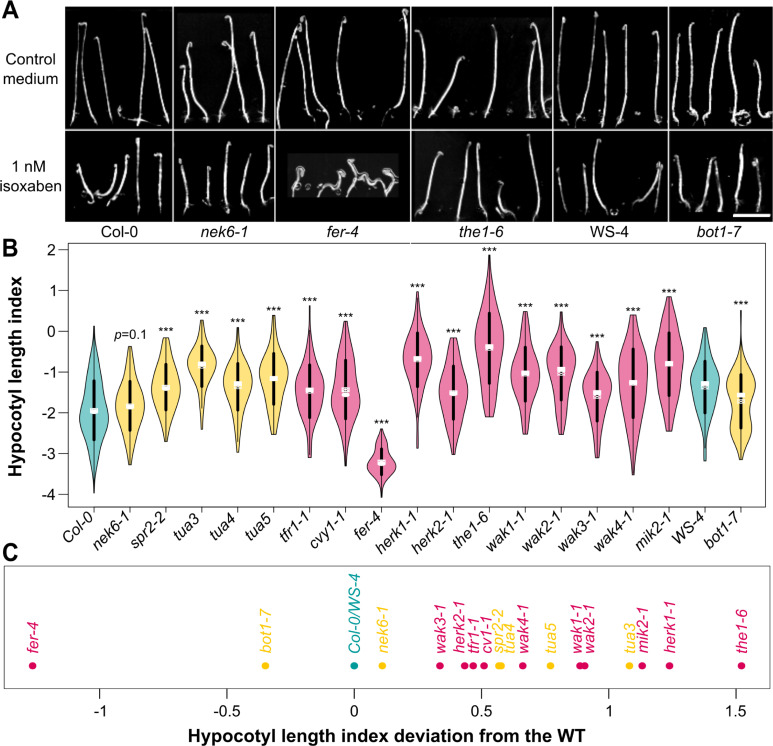
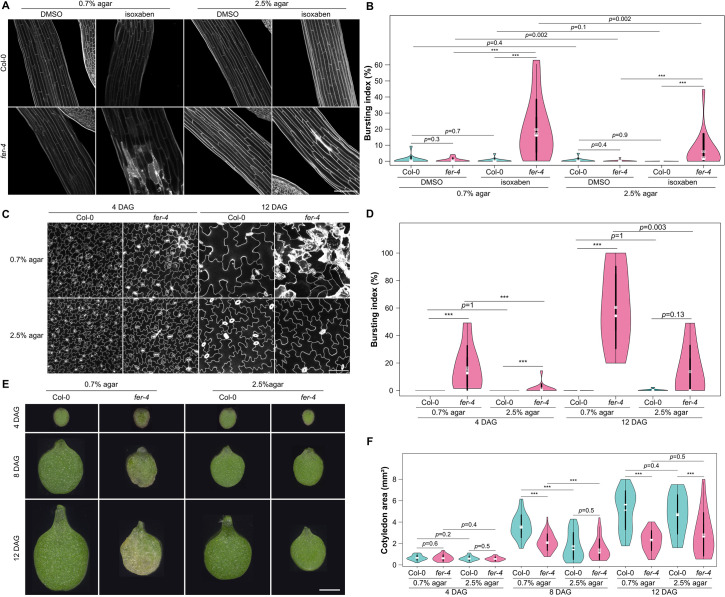
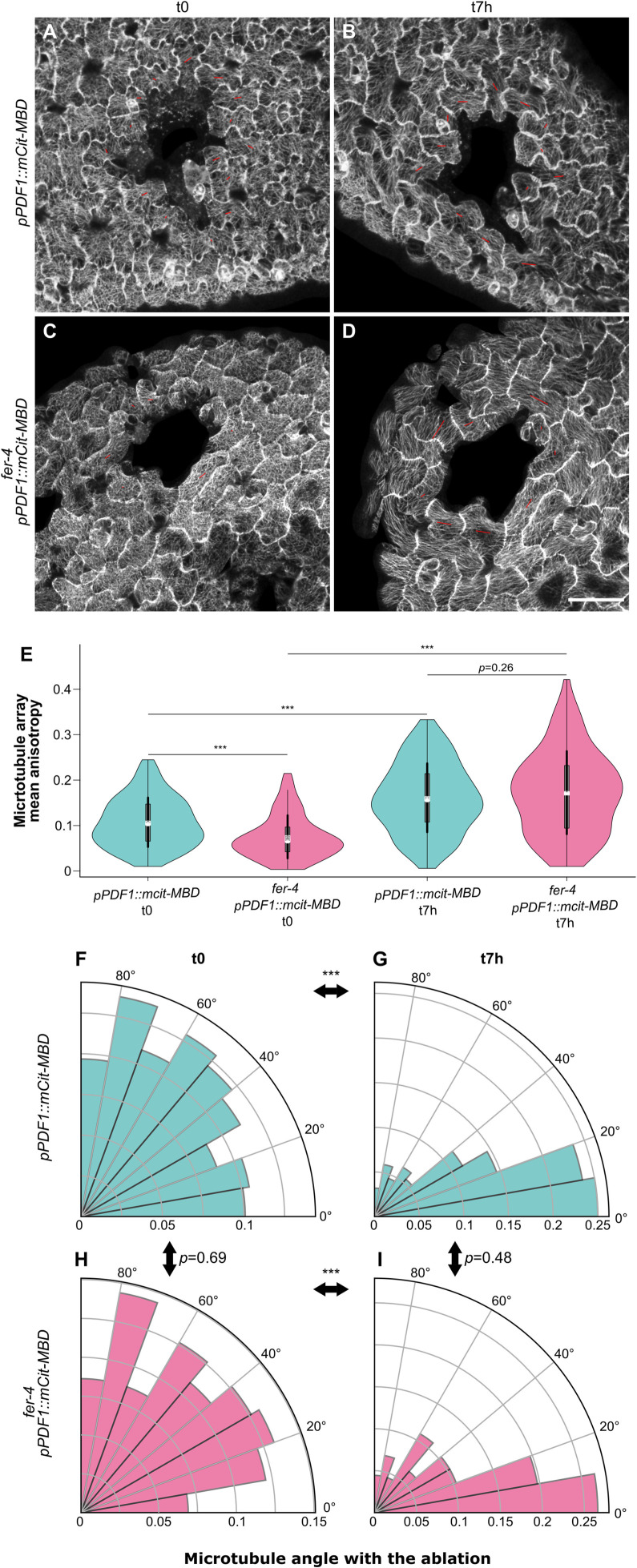
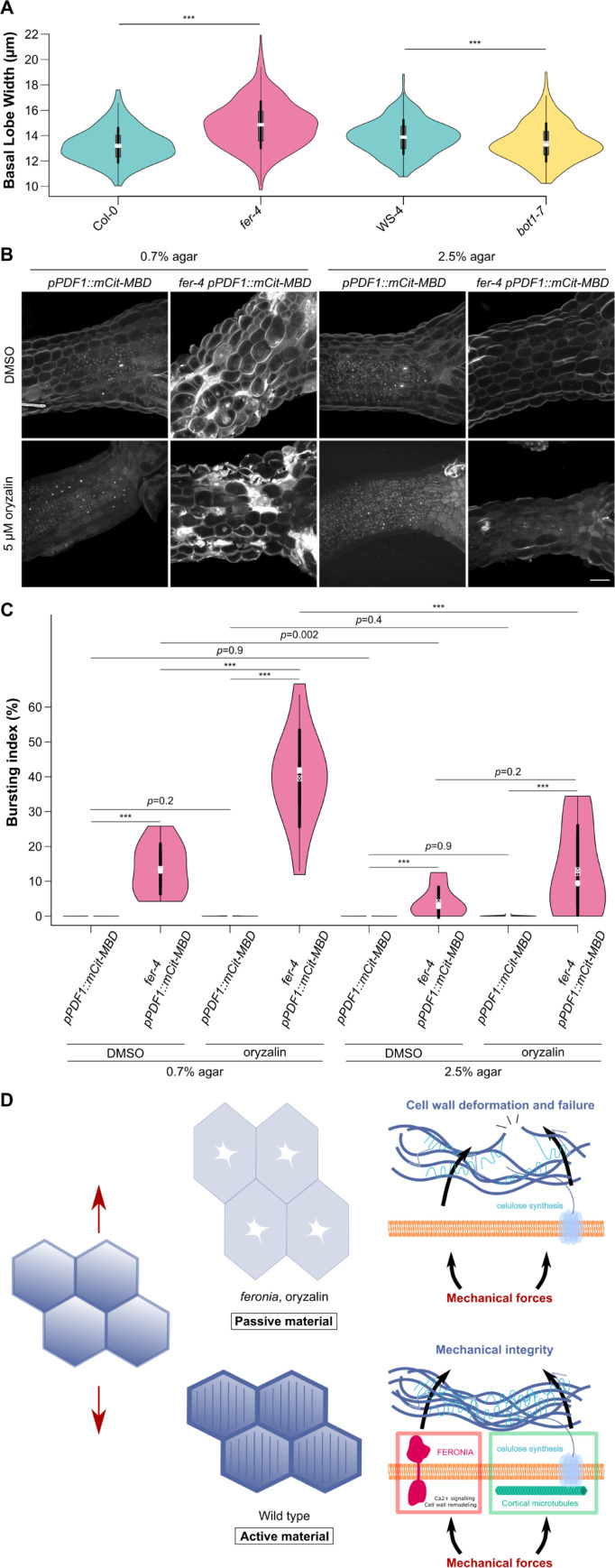
To survive, cells must constantly resist mechanical stress. In plants, this involves the reinforcement of cell walls, notably through microtubule-dependent cellulose deposition. How wall sensing might contribute to this response is unknown. Here, we tested whether the microtubule response to stress acts downstream of known wall sensors. Using a multistep screen with 11 mutant lines, we identify FERONIA (FER) as the primary candidate for the cell's response to stress in the shoot. However, this does not imply that FER acts upstream of the microtubule response to stress. In fact, when performing mechanical perturbations, we instead show that the expected microtubule response to stress does not require FER. We reveal that the feronia phenotype can be partially rescued by reducing tensile stress levels. Conversely, in the absence of both microtubules and FER, cells appear to swell and burst. Altogether, this shows that the microtubule response to stress acts as an independent pathway to resist stress, in parallel to FER. We propose that both pathways are required to maintain the mechanical integrity of plant cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
