

Evaluation of protein kinase D auto-phosphorylation as biomarker for NLRP3 inflammasome activation
- PMID: 34767572
- PMCID: PMC8589197
- DOI: 10.1371/journal.pone.0248668
Evaluation of protein kinase D auto-phosphorylation as biomarker for NLRP3 inflammasome activation
Abstract
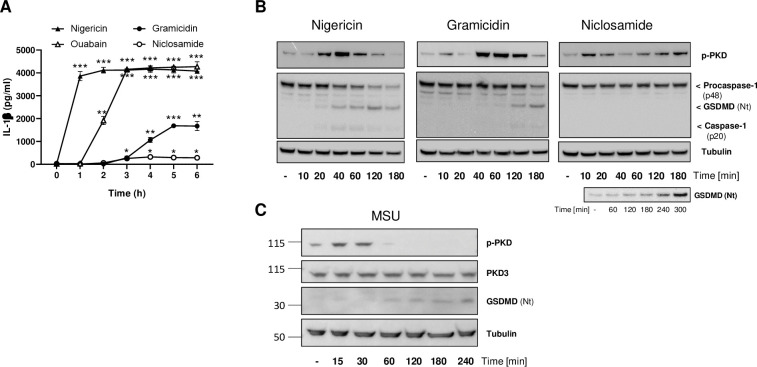
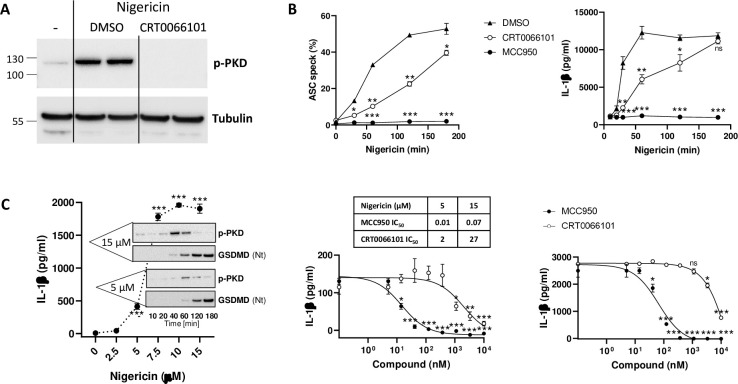
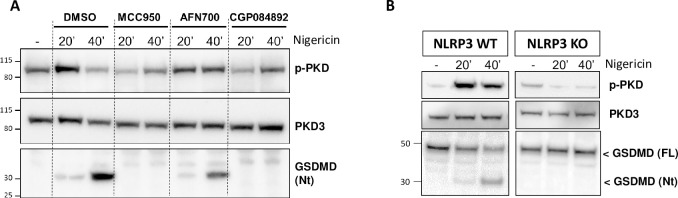
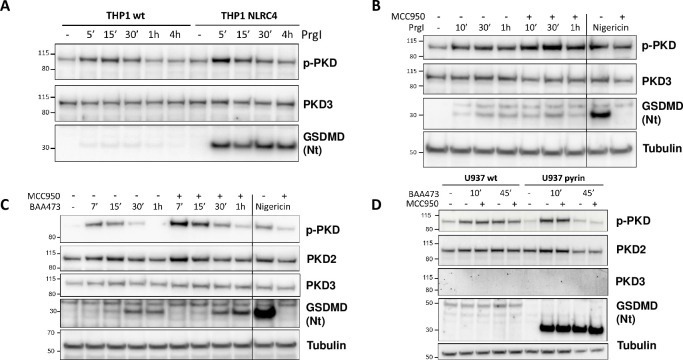
Background: The NLRP3 inflammasome is a critical component of sterile inflammation, which is involved in many diseases. However, there is currently no known proximal biomarker for measuring NLRP3 activation in pathological conditions. Protein kinase D (PKD) has emerged as an important NLRP3 kinase that catalyzes the release of a phosphorylated NLRP3 species that is competent for inflammasome complex assembly.
Methods: To explore the potential for PKD activation to serve as a selective biomarker of the NLRP3 pathway, we tested various stimulatory conditions in THP-1 and U937 cell lines, probing the inflammasome space beyond NLRP3. We analyzed the correlation between PKD activation (monitored by its auto-phosphorylation) and functional inflammasome readouts.
Results: PKD activation/auto-phosphorylation always preceded cleavage of caspase-1 and gasdermin D, and treatment with the PKD inhibitor CRT0066101 could block NLRP3 inflammasome assembly and interleukin-1β production. Conversely, blocking NLRP3 either genetically or using the MCC950 inhibitor prevented PKD auto-phosphorylation, indicating a bidirectional functional crosstalk between NLRP3 and PKD. Further assessments of the pyrin and NLRC4 pathways, however, revealed that PKD auto-phosphorylation can be triggered by a broad range of stimuli unrelated to NLRP3 inflammasome assembly.
Conclusion: Although PKD and NLRP3 become functionally interconnected during NLRP3 activation, the promiscuous reactivity of PKD challenges its potential use for tracing the NLRP3 inflammasome pathway.
Conflict of interest statement
All authors are employees at Novartis Institutes for Biomedical Research (NIBR) at time of studies. This does not alter adherence to PLOS ONE policies on sharing data and materials. None of the authors have competing interest relating to employment,consultancy, patents, products in development, marketed product or else.
Figures
Similar articles
-
Thrombin receptor PAR4 drives canonical NLRP3 inflammasome signaling in the heart.Basic Res Cardiol. 2020 Jan 7;115(2):10. doi: 10.1007/s00395-019-0771-9. Basic Res Cardiol. 2020. PMID: 31912235 Free PMC article.
-
Francisella induced microparticulate caspase-1/gasdermin-D activation is regulated by NLRP3 independent of Pyrin.PLoS One. 2018 Dec 31;13(12):e0209931. doi: 10.1371/journal.pone.0209931. eCollection 2018. PLoS One. 2018. PMID: 30596757 Free PMC article.
-
Pharmacological inhibition of IKKβ dampens NLRP3 inflammasome activation after priming in the human myeloid cell line THP-1.Biochem Biophys Res Commun. 2021 Mar 19;545:177-182. doi: 10.1016/j.bbrc.2021.01.051. Epub 2021 Feb 6. Biochem Biophys Res Commun. 2021. PMID: 33561652
-
Regulation of the NLRP3 Inflammasome by Posttranslational Modifications.J Immunol. 2022 Jan 15;208(2):286-292. doi: 10.4049/jimmunol.2100734. J Immunol. 2022. PMID: 35017218 Review.
-
Effects of phosphorylation on the NLRP3 inflammasome.Arch Biochem Biophys. 2019 Jul 30;670:43-57. doi: 10.1016/j.abb.2019.02.020. Epub 2019 Mar 5. Arch Biochem Biophys. 2019. PMID: 30844378 Review.
Cited by
-
PHOrming the inflammasome: phosphorylation is a critical switch in inflammasome signalling.Biochem Soc Trans. 2021 Dec 17;49(6):2495-2507. doi: 10.1042/BST20200987. Biochem Soc Trans. 2021. PMID: 34854899 Free PMC article. Review.
-
A phospho-harmonic orchestra plays the NLRP3 score.Front Immunol. 2023 Nov 3;14:1281607. doi: 10.3389/fimmu.2023.1281607. eCollection 2023. Front Immunol. 2023. PMID: 38022631 Free PMC article. Review.
-
Interleukin 38 alleviates aortic valve calcification by inhibition of NLRP3.Proc Natl Acad Sci U S A. 2022 Sep 6;119(36):e2202577119. doi: 10.1073/pnas.2202577119. Epub 2022 Aug 29. Proc Natl Acad Sci U S A. 2022. PMID: 36037361 Free PMC article.
References
-
- Schroder K, Sagulenko V, Zamoshnikova A, Richards AA, Cridland JA, Irvine KM, et al.. Acute lipopolysaccharide priming boosts inflammasome activation independently of inflammasome sensor induction. Immunobiology [Internet]. 2012;217(12):1325–9. Available from: doi: 10.1016/j.imbio.2012.07.020 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
