

MTHFD2 is a metabolic checkpoint controlling effector and regulatory T cell fate and function
- PMID: 34767747
- PMCID: PMC8755618
- DOI: 10.1016/j.immuni.2021.10.011
MTHFD2 is a metabolic checkpoint controlling effector and regulatory T cell fate and function
Abstract
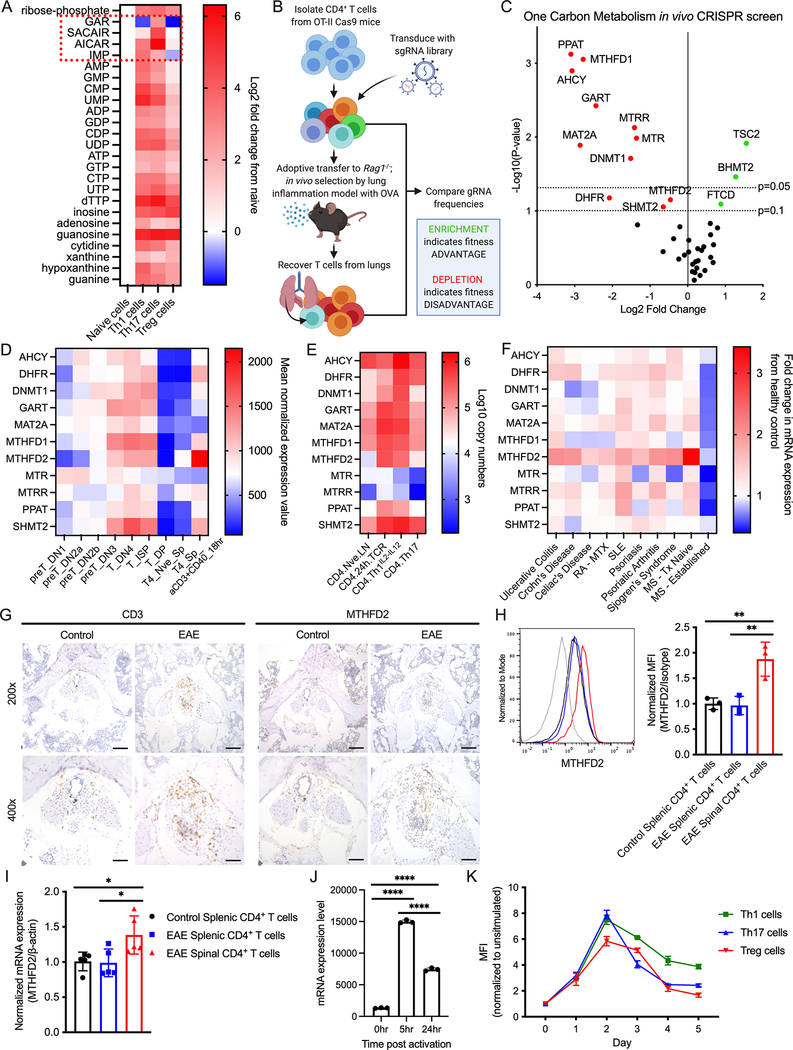
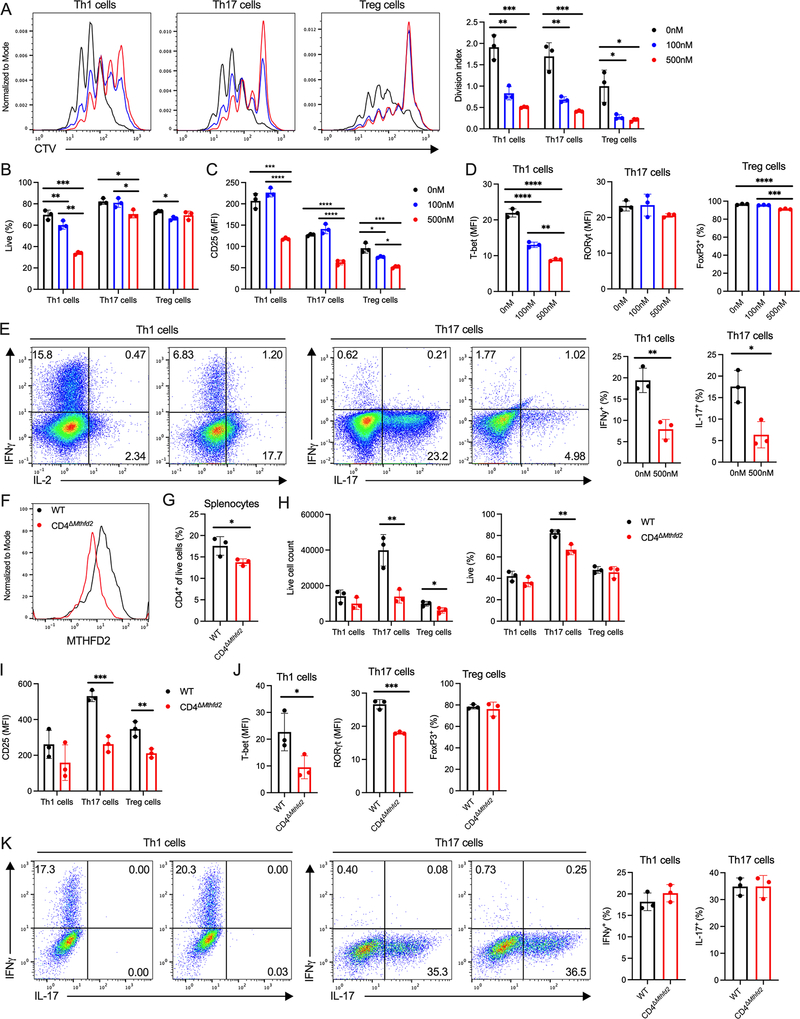
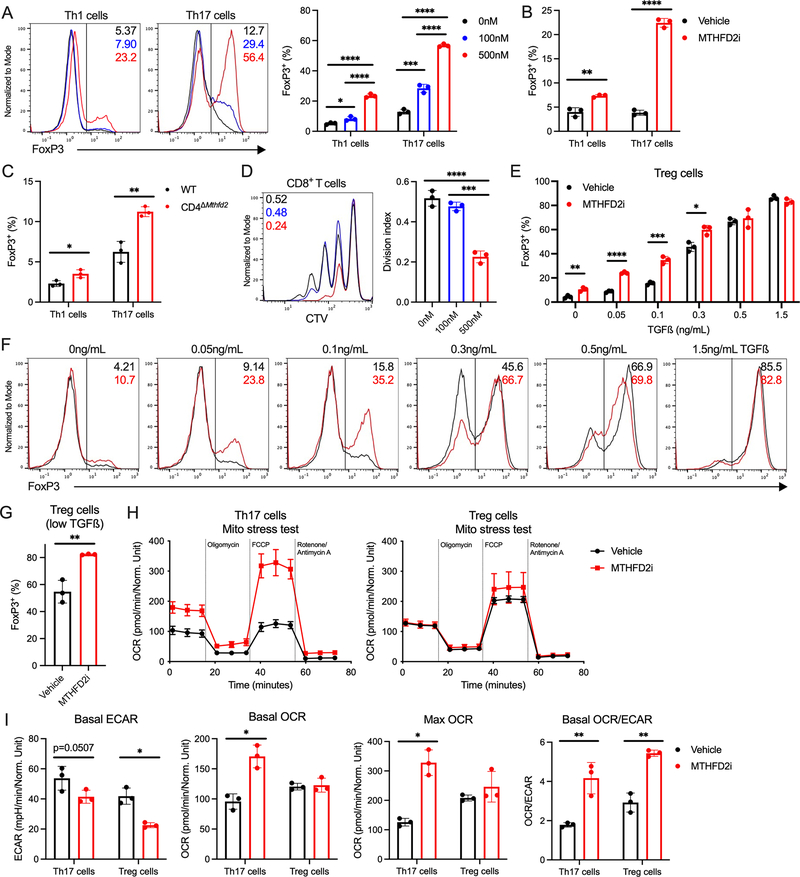
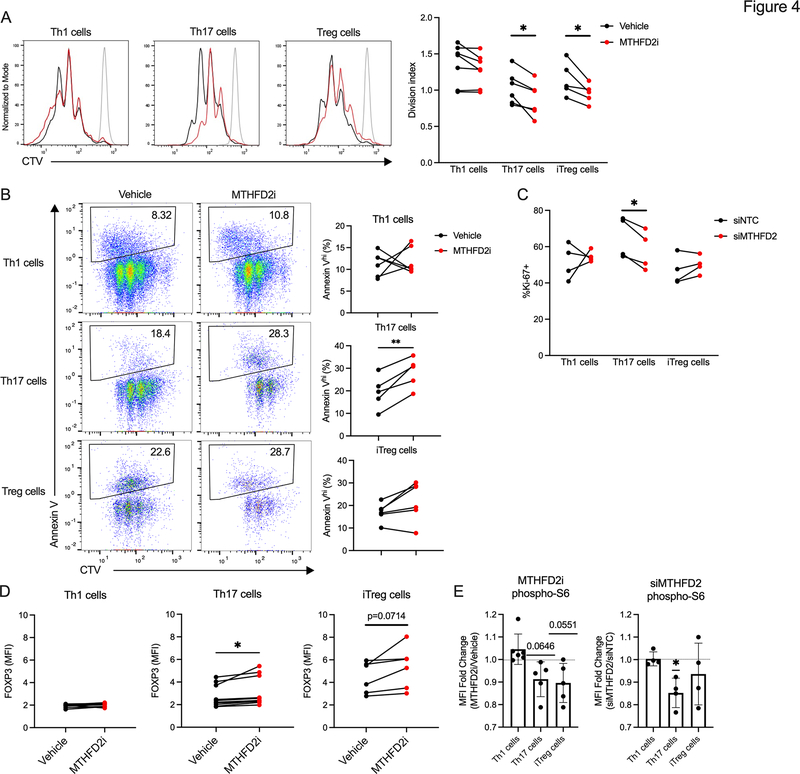
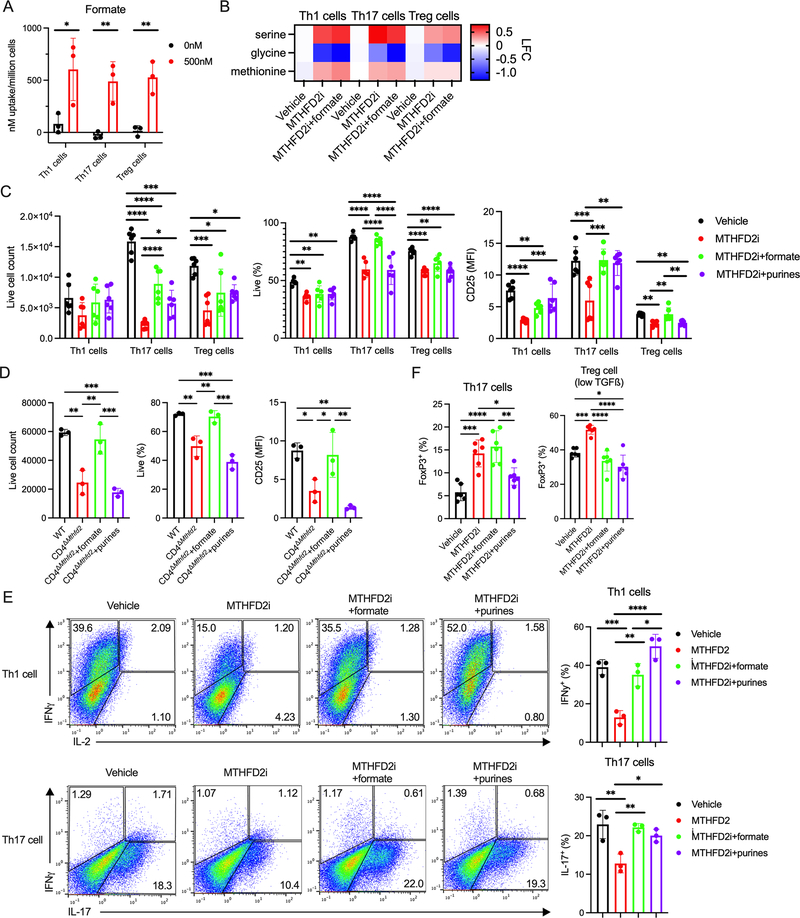
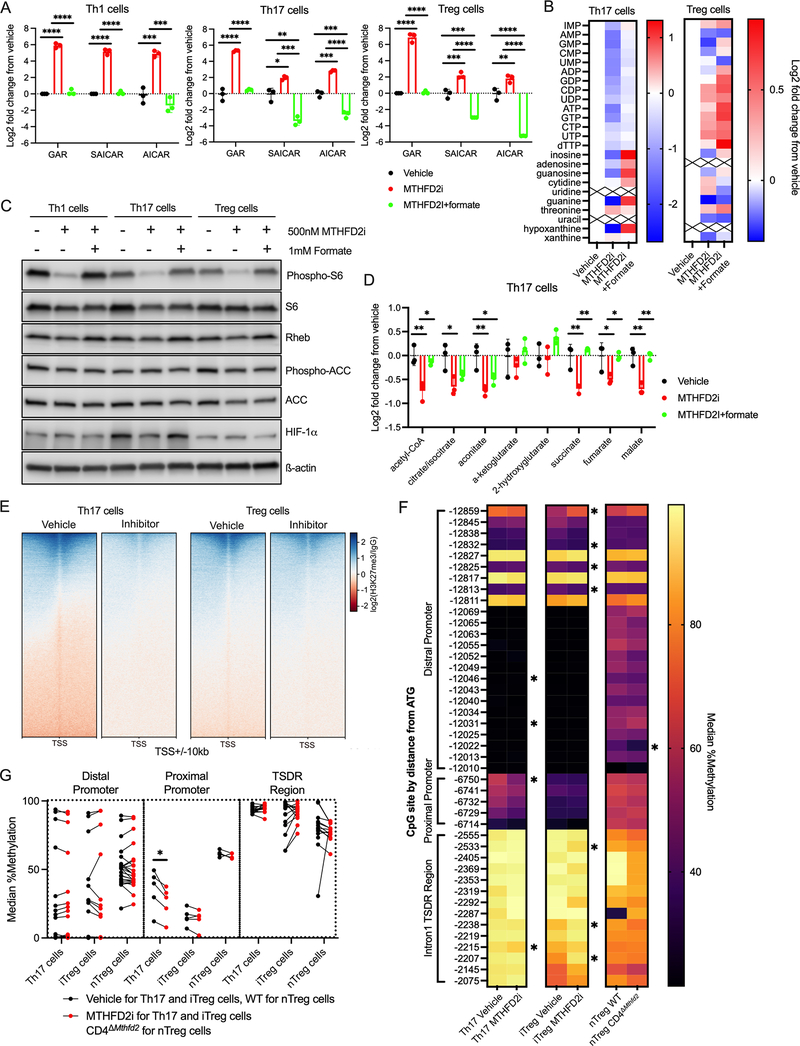
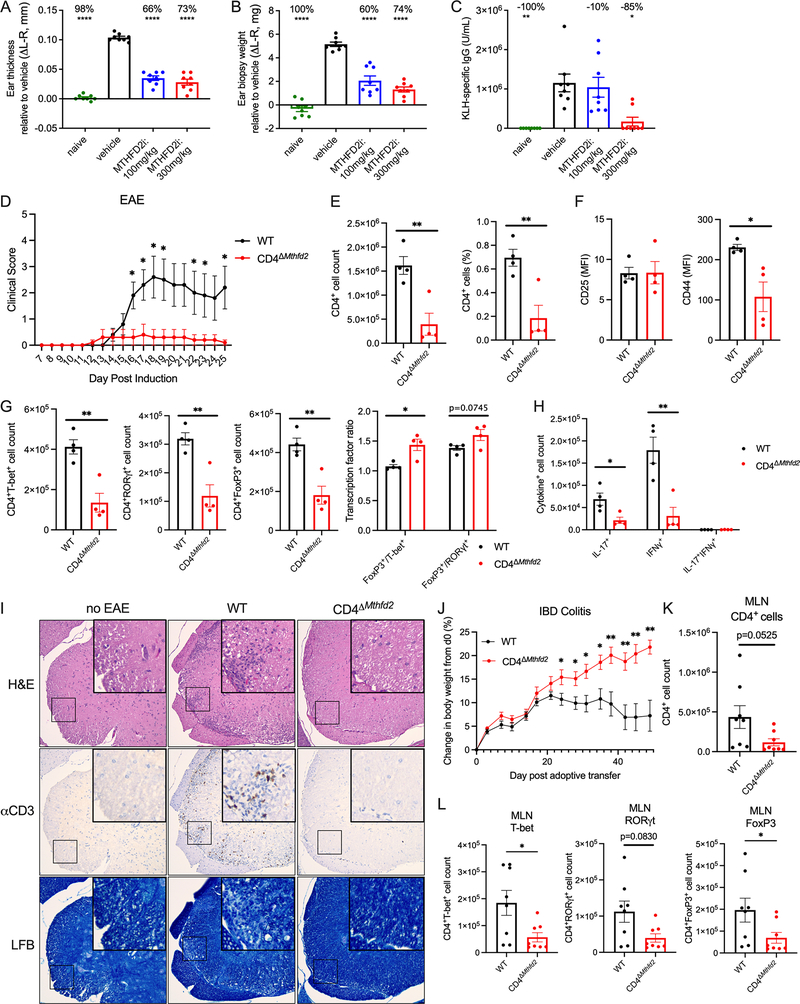
Antigenic stimulation promotes T cell metabolic reprogramming to meet increased biosynthetic, bioenergetic, and signaling demands. We show that the one-carbon (1C) metabolism enzyme methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) regulates de novo purine synthesis and signaling in activated T cells to promote proliferation and inflammatory cytokine production. In pathogenic T helper-17 (Th17) cells, MTHFD2 prevented aberrant upregulation of the transcription factor FoxP3 along with inappropriate gain of suppressive capacity. MTHFD2 deficiency also promoted regulatory T (Treg) cell differentiation. Mechanistically, MTHFD2 inhibition led to depletion of purine pools, accumulation of purine biosynthetic intermediates, and decreased nutrient sensor mTORC1 signaling. MTHFD2 was also critical to regulate DNA and histone methylation in Th17 cells. Importantly, MTHFD2 deficiency reduced disease severity in multiple in vivo inflammatory disease models. MTHFD2 is thus a metabolic checkpoint to integrate purine metabolism with pathogenic effector cell signaling and is a potential therapeutic target within 1C metabolism pathways.
Keywords: CD4(+) T cells; CRISPR screen; MTHFD2; T cell differentiation; inflammation; mTORC1; metabolic checkpoint; methylation; one carbon metabolism; purine metabolism.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.C.R. is a founder, scientific advisory board member, and stockholder of Sitryx Therapeutics; a scientific advisory board member and stockholder of Caribou Biosciences; a member of the scientific advisory board of Nirogy Therapeutics; has consulted for Merck, Pfizer, and Mitobridge within the past 3 years; and has received research support from Incyte Corp., Calithera Biosciences, and Tempest Therapeutics. J.D.R. is a co-founder and stockholder in Raze Therapeutics, Toran, Serien Therapeutics, and Farber Partners and an advisor and stockholder in Agios Pharmaceuticals, Kadmon Pharmaceuticals, Bantam Pharmaceuticals, Colorado Research Partners, Rafael Holdings, the Barer Institute, and L.E.A.F. Pharmaceuticals; he has received consulting fees and research funding from Pfizer and Rafael and is the inventor of patents held by Princeton University. A.M.C., P.F., and T.B. are employees of Sitryx Therapeutics.
Figures
Comment in
-
The folate way to T cell fate.Immunity. 2022 Jan 11;55(1):1-3. doi: 10.1016/j.immuni.2021.12.009. Immunity. 2022. PMID: 35021051
-
MTHFD2, metabolic mastermind of CD4+ T cell destiny.Sci Immunol. 2022 Mar 4;7(69):eabo6765. doi: 10.1126/sciimmunol.abo6765. Epub 2022 Mar 4. Sci Immunol. 2022. PMID: 35245088
References
-
- Bantug GR, Galluzzi L, Kroemer G, and Hess C (2018). The spectrum of T cell metabolism in health and disease. Nat Rev Immunol 18, 19–34. - PubMed
-
- Brown PM, Pratt AG, and Isaacs JD (2016). Mechanism of action of methotrexate in rheumatoid arthritis, and the search for biomarkers. Nat Rev Rheumatol 12, 731–742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10 OD023475/OD/NIH HHS/United States
- R01 HL118979/HL/NHLBI NIH HHS/United States
- R01 DK105550/DK/NIDDK NIH HHS/United States
- T32 DK101003/DK/NIDDK NIH HHS/United States
- F30 CA239367/CA/NCI NIH HHS/United States
- T32 GM007347/GM/NIGMS NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- T32 GM152284/GM/NIGMS NIH HHS/United States
- R01 HL136664/HL/NHLBI NIH HHS/United States
- R01 CA217987/CA/NCI NIH HHS/United States
- R01 AI137075/AI/NIAID NIH HHS/United States
- P30 CA086862/CA/NCI NIH HHS/United States
- R01 AI153167/AI/NIAID NIH HHS/United States
- R01 HL122554/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
