

Mitochondria as a Cellular Hub in Infection and Inflammation
- PMID: 34768767
- PMCID: PMC8583510
- DOI: 10.3390/ijms222111338
Mitochondria as a Cellular Hub in Infection and Inflammation
Abstract
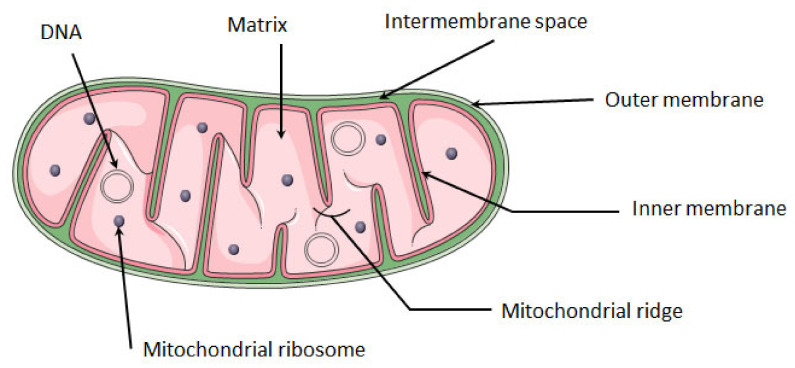
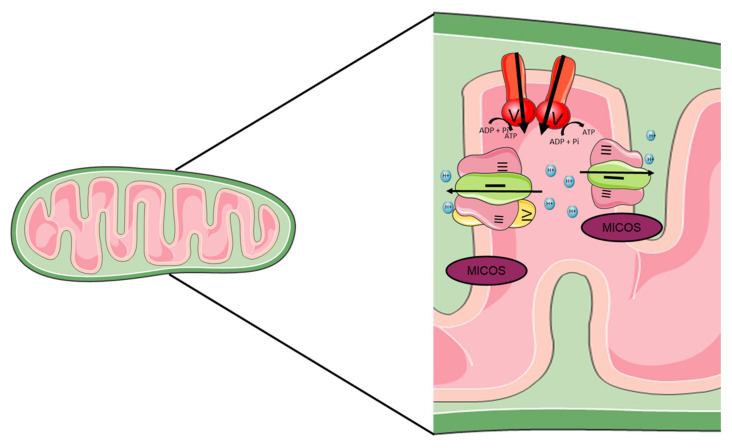
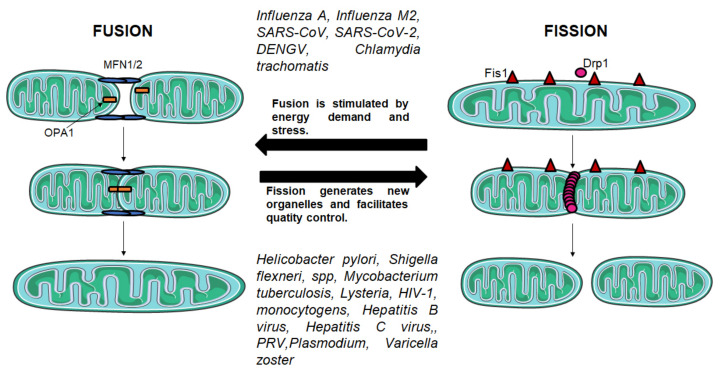
Mitochondria are the energy center of the cell. They are found in the cell cytoplasm as dynamic networks where they adapt energy production based on the cell's needs. They are also at the center of the proinflammatory response and have essential roles in the response against pathogenic infections. Mitochondria are a major site for production of Reactive Oxygen Species (ROS; or free radicals), which are essential to fight infection. However, excessive and uncontrolled production can become deleterious to the cell, leading to mitochondrial and tissue damage. Pathogens exploit the role of mitochondria during infection by affecting the oxidative phosphorylation mechanism (OXPHOS), mitochondrial network and disrupting the communication between the nucleus and the mitochondria. The role of mitochondria in these biological processes makes these organelle good targets for the development of therapeutic strategies. In this review, we presented a summary of the endosymbiotic origin of mitochondria and their involvement in the pathogen response, as well as the potential promising mitochondrial targets for the fight against infectious diseases and chronic inflammatory diseases.
Keywords: infection; infection disease; inflammation; inflammatory disease; mitochondria; mitochondria dysfunction; mitochondrial bioenergetics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
Publication types
MeSH terms
Grants and funding
- none/the Institut National de la Santé et de la Recherche Médicale (INSERM)
- AMIDEX "International_2018" MITOMUTCHAGAS/the Aix-Marseille University
- Br-Fr-Chagas/the French Agency for Research (Agence Nationale de la Recherche-ANR)
- landscardio/the French Agency for Research (Agence Nationale de la Recherche-ANR)
- none/the CNPq (Brazilian Council for Scientific and Technological Development)
- 2013/50302-3/the FAPESP (São Paulo State Research Funding Agency Brazil)
- 2014/50890-5/the FAPESP (São Paulo State Research Funding Agency Brazil)
- 2 P50 AI098461-02/the National Institutes of Health/USA
- 2U19AI098461-06/the National Institutes of Health/USA
- none/the Inserm Cross-Cutting Project GOLD
- Institute MarMaRa AMX-19-IET-007/the Excellence Initiative of Aix-Marseille University - A*Midex
LinkOut - more resources
Full Text Sources
