

Small RNA F6 Provides Mycobacterium smegmatis Entry into Dormancy
- PMID: 34768965
- PMCID: PMC8583896
- DOI: 10.3390/ijms222111536
Small RNA F6 Provides Mycobacterium smegmatis Entry into Dormancy
Abstract
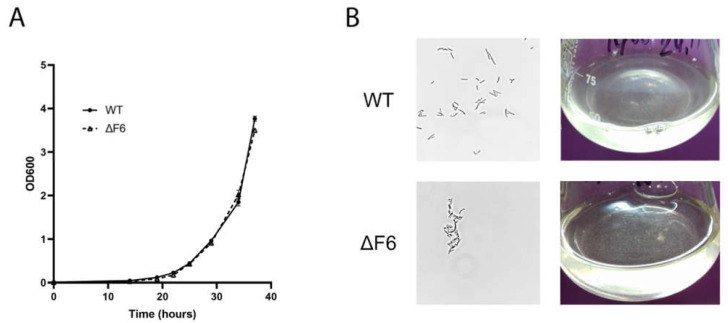
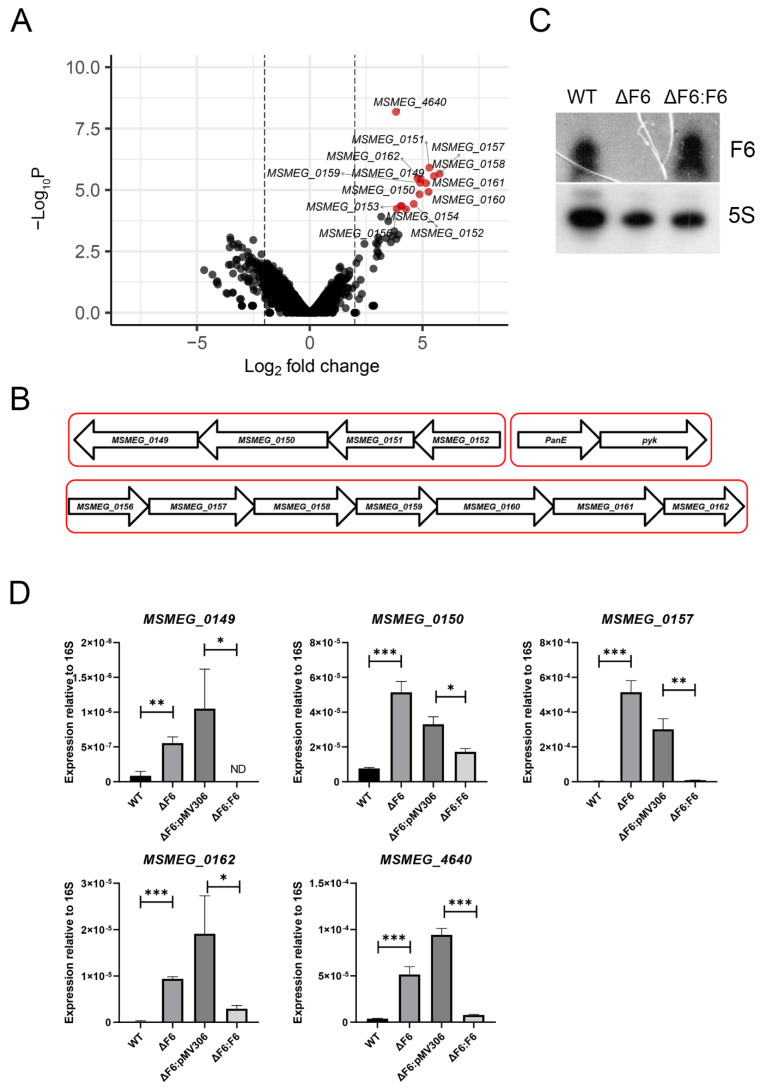
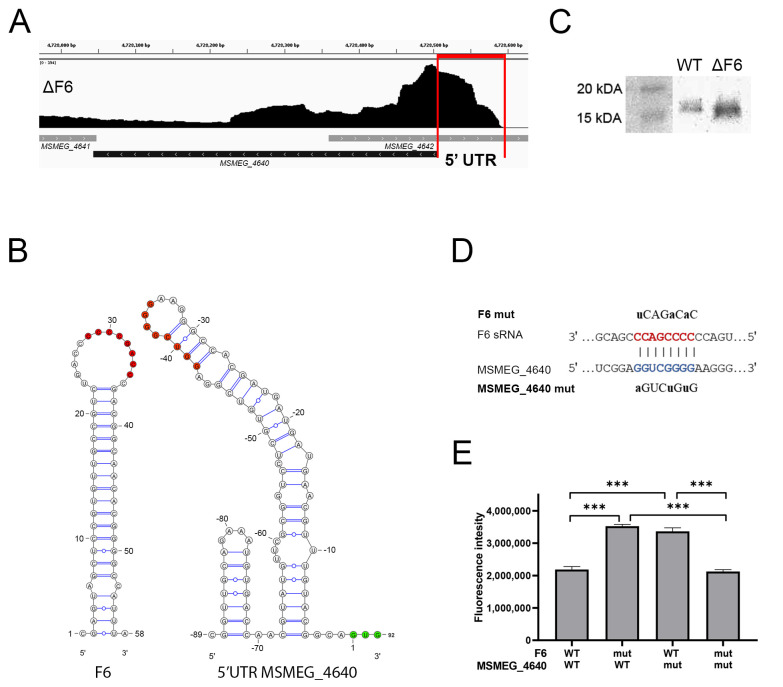
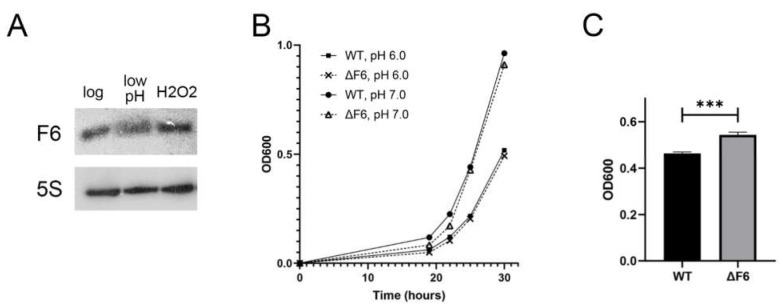
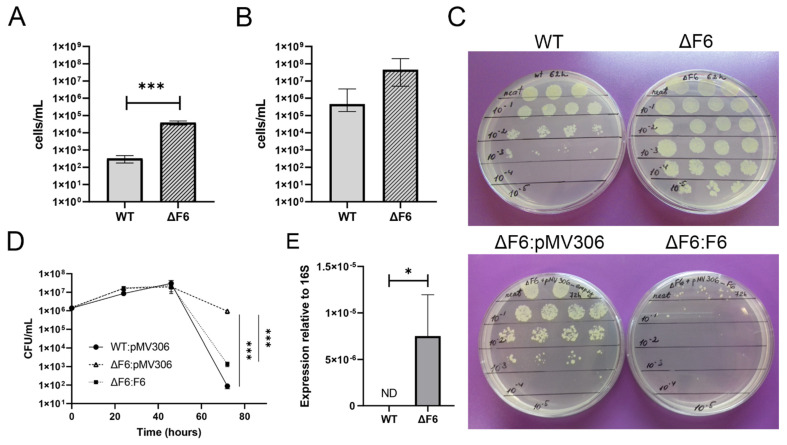
Regulatory small non-coding RNAs play a significant role in bacterial adaptation to changing environmental conditions. Various stresses such as hypoxia and nutrient starvation cause a reduction in the metabolic activity of Mycobacterium smegmatis, leading to entry into dormancy. We investigated the functional role of F6, a small RNA of M. smegmatis, and constructed an F6 deletion strain of M. smegmatis. Using the RNA-seq approach, we demonstrated that gene expression changes that accompany F6 deletion contributed to bacterial resistance against oxidative stress. We also found that F6 directly interacted with 5'-UTR of MSMEG_4640 mRNA encoding RpfE2, a resuscitation-promoting factor, which led to the downregulation of RpfE2 expression. The F6 deletion strain was characterized by the reduced ability to enter into dormancy (non-culturability) in the potassium deficiency model compared to the wild-type strain, indicating that F6 significantly contributes to bacterial adaptation to non-optimal growth conditions.
Keywords: F6; Mycobacterium smegmatis; adaptation to stresses; dormancy; non-culturability; resuscitation promoting factor RpfE2; small non-coding RNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Gerrick E.R., Barbier T., Chase M.R., Xu R., François J., Lin V.H., Szucs M.J., Rock J.M., Ahmad R., Tjaden B. Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. USA. 2018;115:6464–6469. doi: 10.1073/pnas.1718003115. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
