

Establishment of Human-Induced Pluripotent Stem Cell-Derived Neurons-A Promising In Vitro Model for a Molecular Study of Rabies Virus and Host Interaction
- PMID: 34769416
- PMCID: PMC8584829
- DOI: 10.3390/ijms222111986
Establishment of Human-Induced Pluripotent Stem Cell-Derived Neurons-A Promising In Vitro Model for a Molecular Study of Rabies Virus and Host Interaction
Abstract
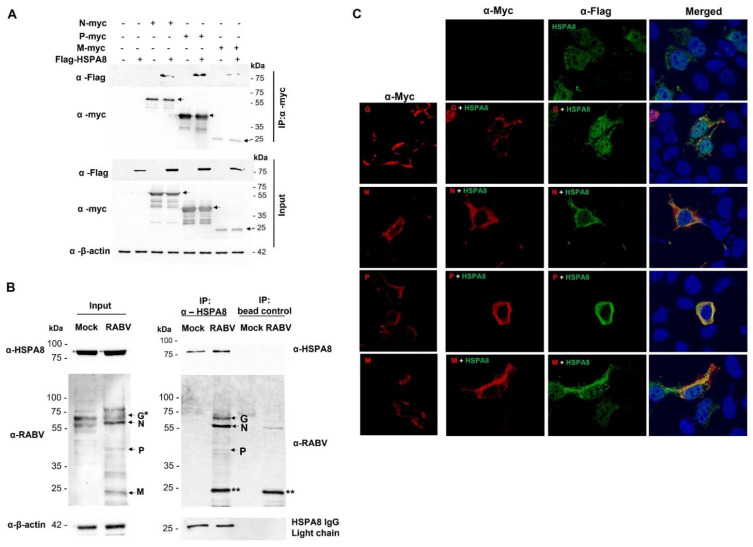
Rabies is a deadly viral disease caused by the rabies virus (RABV), transmitted through a bite of an infected host, resulting in irreversible neurological symptoms and a 100% fatality rate in humans. Despite many aspects describing rabies neuropathogenesis, numerous hypotheses remain unanswered and concealed. Observations obtained from infected primary neurons or mouse brain samples are more relevant to human clinical rabies than permissive cell lines; however, limitations regarding the ethical issue and sample accessibility become a hurdle for discovering new insights into virus-host interplays. To better understand RABV pathogenesis in humans, we generated human-induced pluripotent stem cell (hiPSC)-derived neurons to offer the opportunity for an inimitable study of RABV infection at a molecular level in a pathologically relevant cell type. This study describes the characteristics and detailed proteomic changes of hiPSC-derived neurons in response to RABV infection using LC-MS/MS quantitative analysis. Gene ontology (GO) enrichment of differentially expressed proteins (DEPs) reveals temporal changes of proteins related to metabolic process, immune response, neurotransmitter transport/synaptic vesicle cycle, cytoskeleton organization, and cell stress response, demonstrating fundamental underlying mechanisms of neuropathogenesis in a time-course dependence. Lastly, we highlighted plausible functions of heat shock cognate protein 70 (HSC70 or HSPA8) that might play a pivotal role in regulating RABV replication and pathogenesis. Our findings acquired from this hiPSC-derived neuron platform help to define novel cellular mechanisms during RABV infection, which could be applicable to further studies to widen views of RABV-host interaction.
Keywords: human-induced pluripotent stem cells; in vitro model; neurons; proteomics analysis; rabies virus; virus–host interaction.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
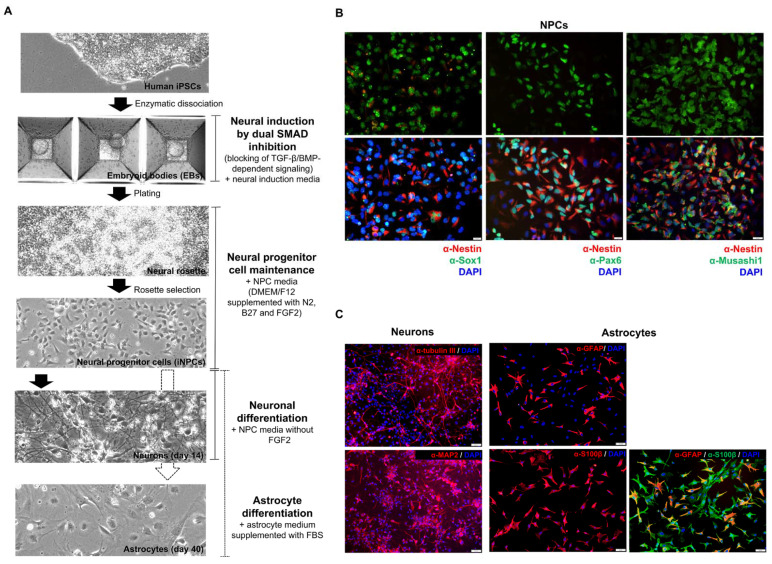
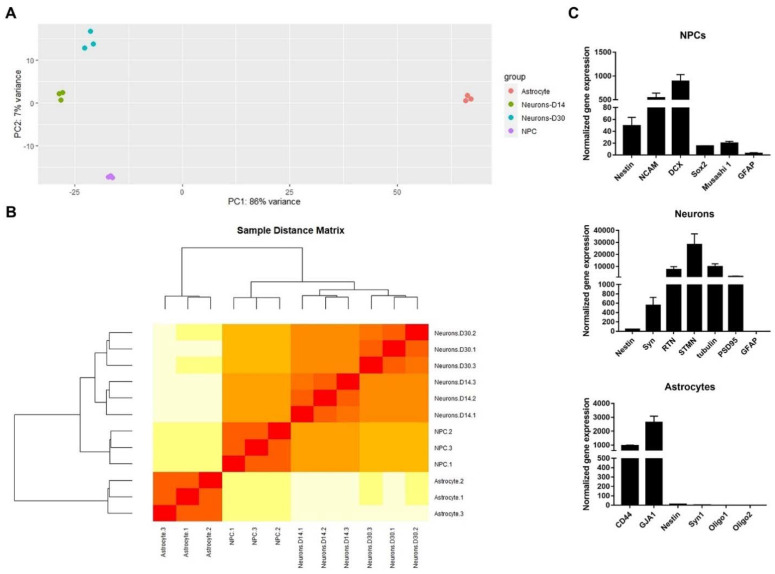
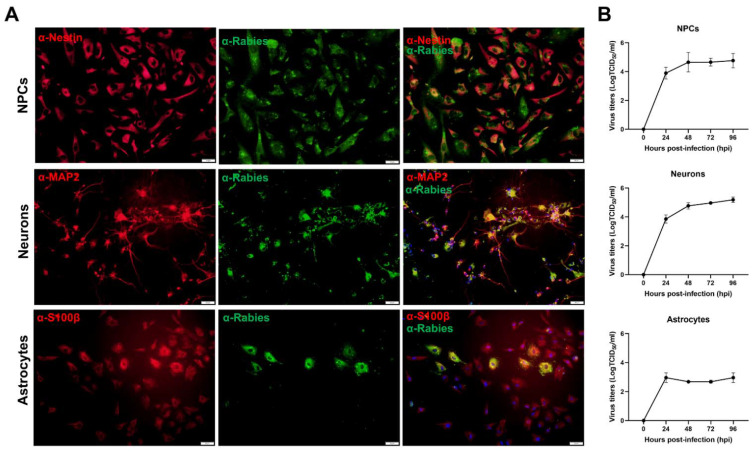
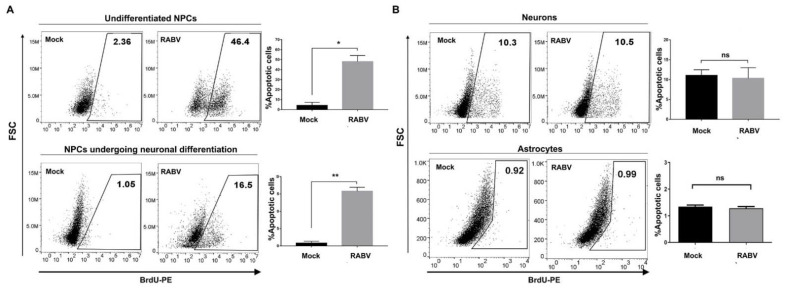
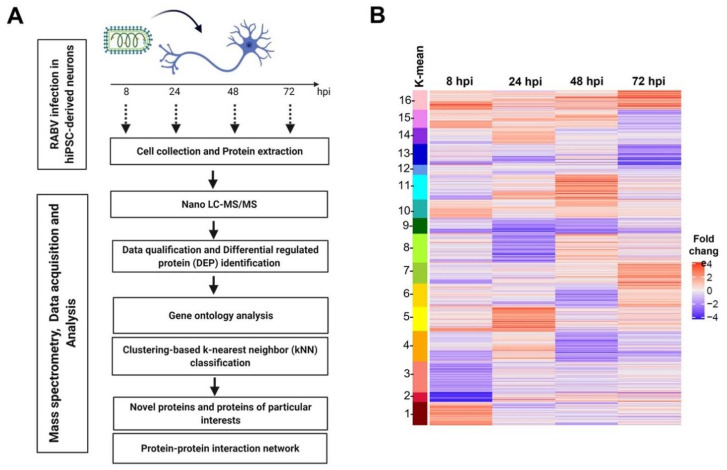
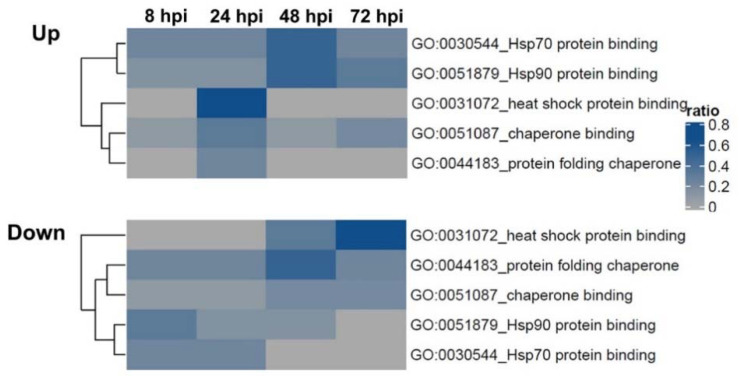
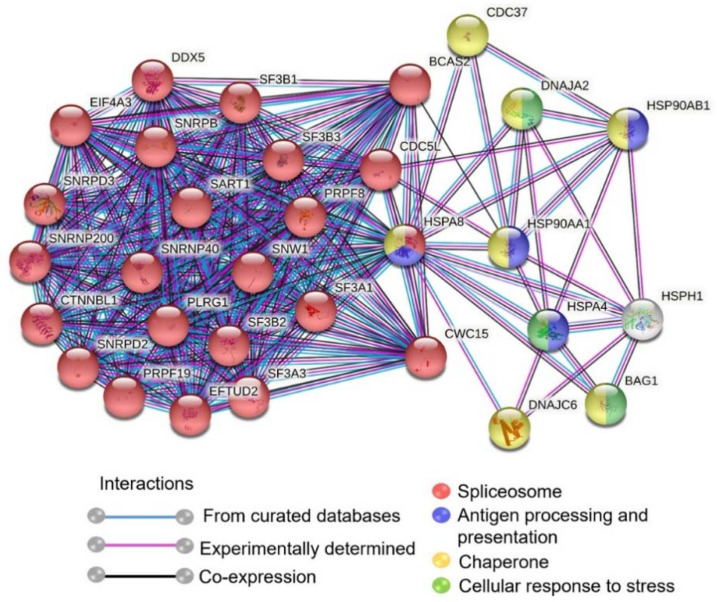
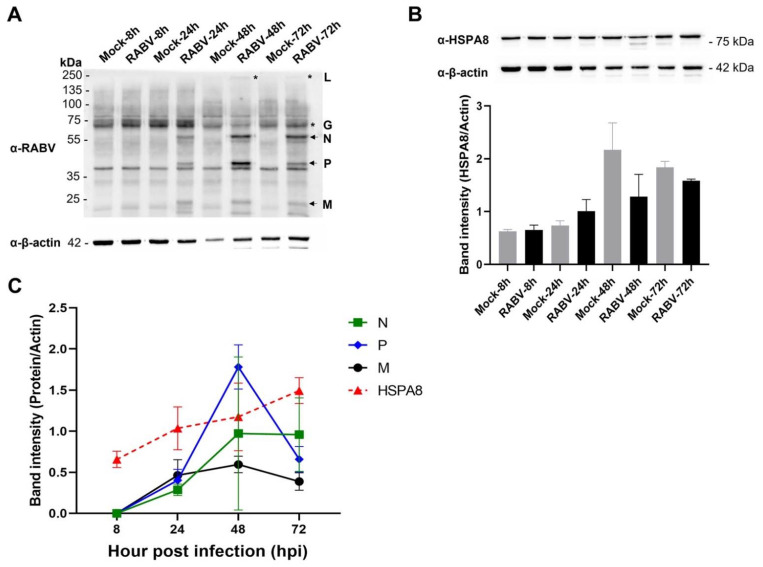
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
