Dynamic mechanochemical feedback between curved membranes and BAR protein self-organization
- PMID: 34772909
- PMCID: PMC8589976
- DOI: 10.1038/s41467-021-26591-3
Dynamic mechanochemical feedback between curved membranes and BAR protein self-organization
Abstract
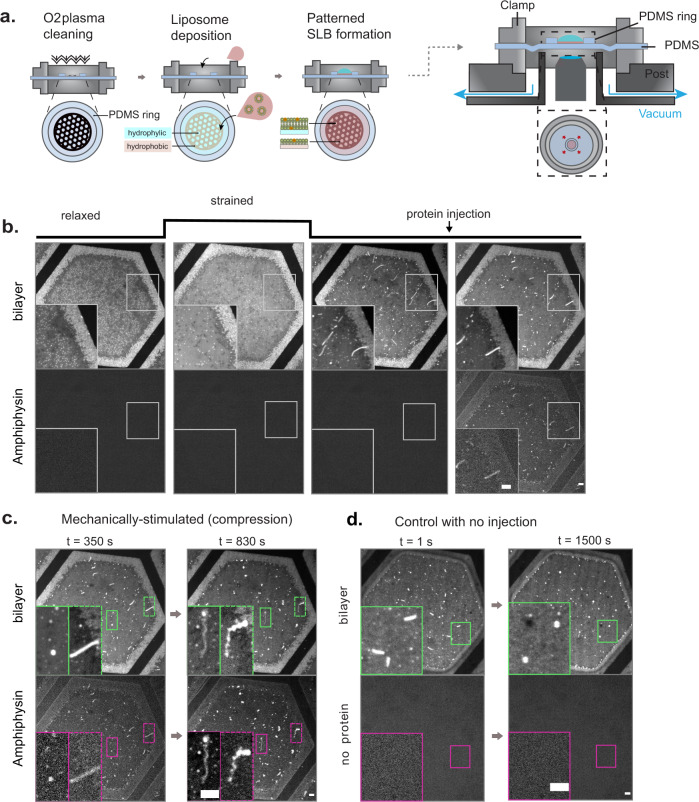
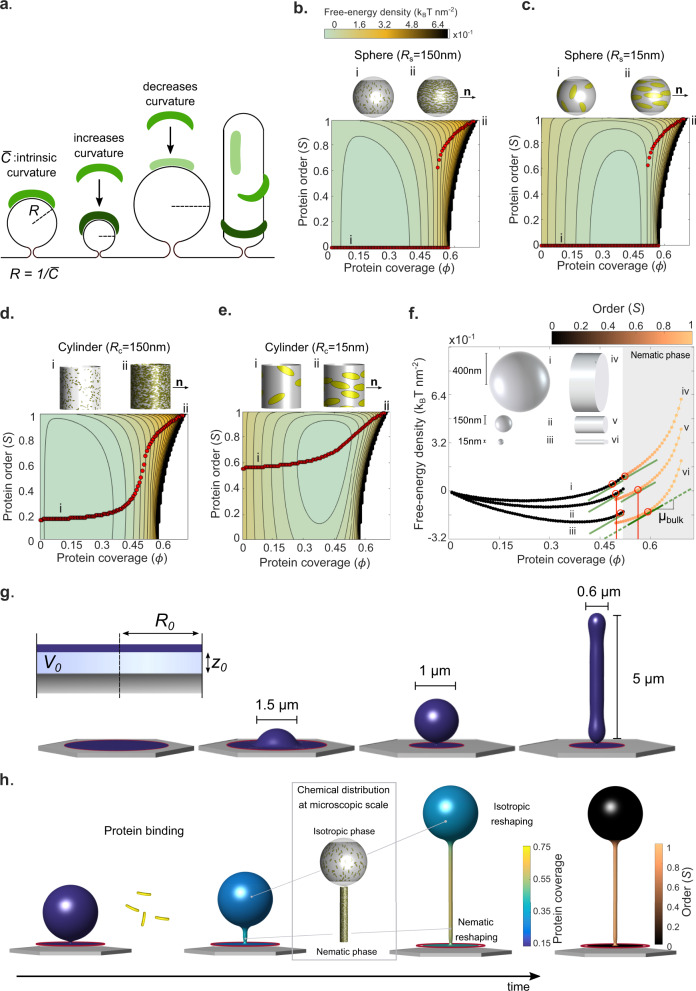
In many physiological situations, BAR proteins reshape membranes with pre-existing curvature (templates), contributing to essential cellular processes. However, the mechanism and the biological implications of this reshaping process remain unclear. Here we show, both experimentally and through modelling, that BAR proteins reshape low curvature membrane templates through a mechanochemical phase transition. This phenomenon depends on initial template shape and involves the co-existence and progressive transition between distinct local states in terms of molecular organization (protein arrangement and density) and membrane shape (template size and spherical versus cylindrical curvature). Further, we demonstrate in cells that this phenomenon enables a mechanotransduction mode, in which cellular stretch leads to the mechanical formation of membrane templates, which are then reshaped into tubules by BAR proteins. Our results demonstrate the interplay between membrane mechanics and BAR protein molecular organization, integrating curvature sensing and generation in a comprehensive framework with implications for cell mechanical responses.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures