

Sensory Adaptation in the Whisker-Mediated Tactile System: Physiology, Theory, and Function
- PMID: 34776857
- PMCID: PMC8586522
- DOI: 10.3389/fnins.2021.770011
Sensory Adaptation in the Whisker-Mediated Tactile System: Physiology, Theory, and Function
Abstract
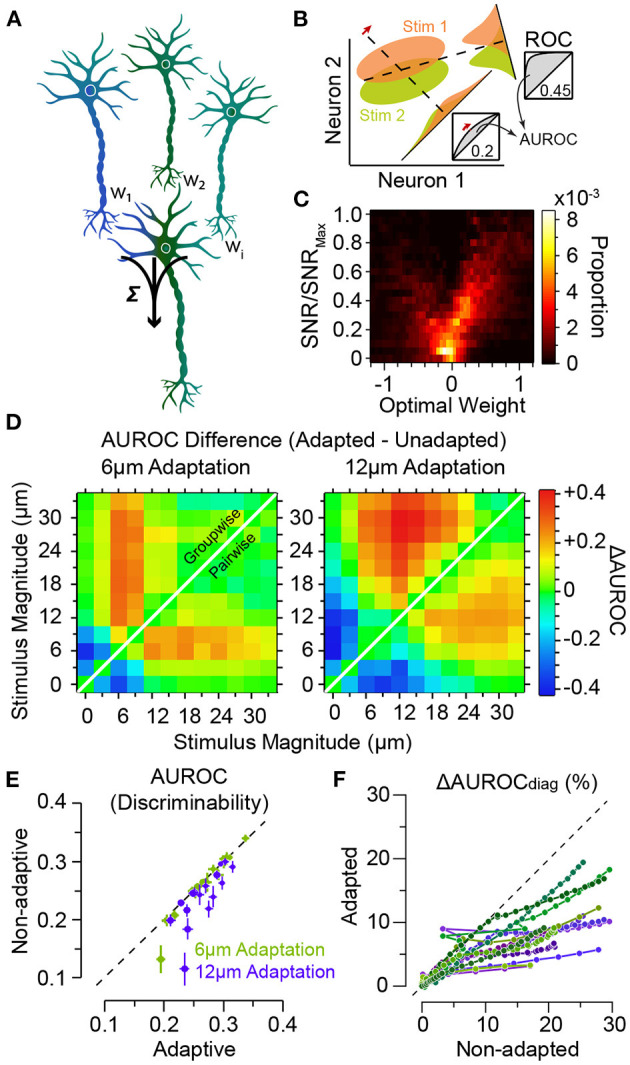
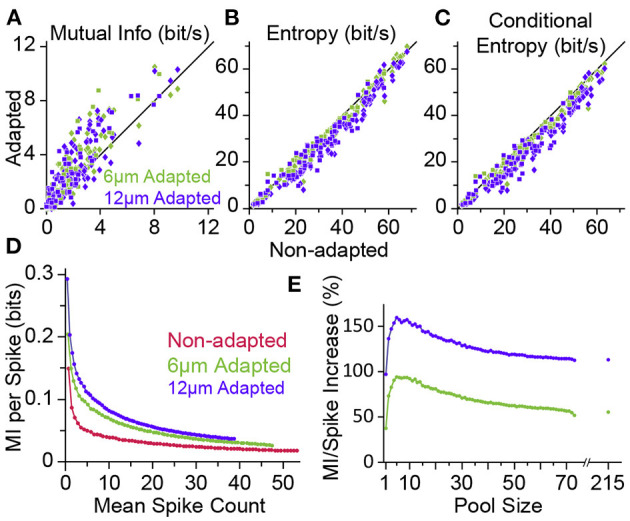
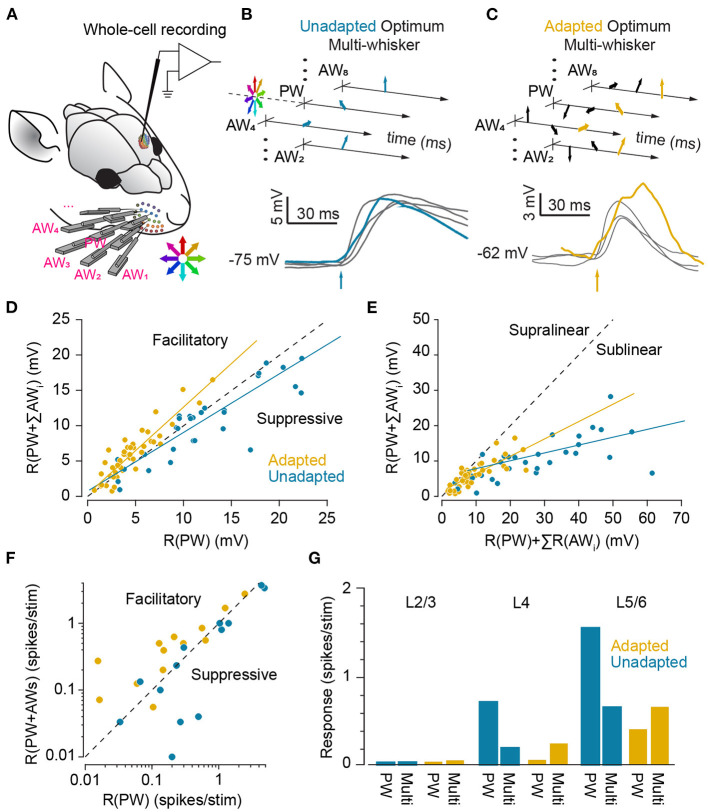
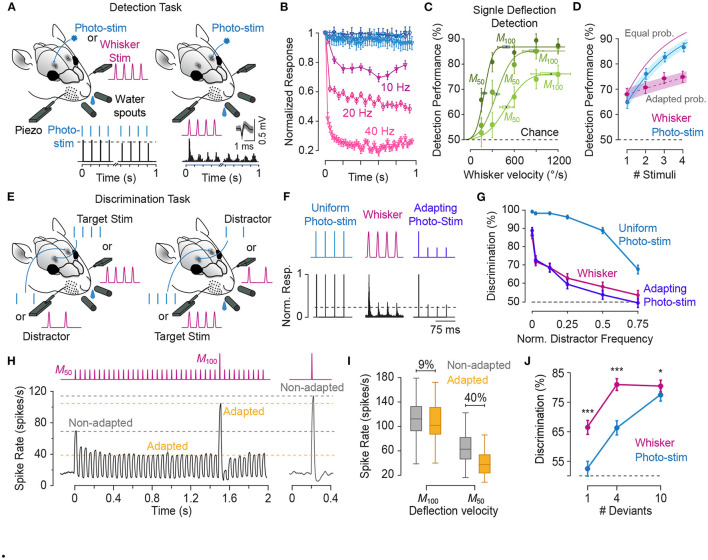
In the natural environment, organisms are constantly exposed to a continuous stream of sensory input. The dynamics of sensory input changes with organism's behaviour and environmental context. The contextual variations may induce >100-fold change in the parameters of the stimulation that an animal experiences. Thus, it is vital for the organism to adapt to the new diet of stimulation. The response properties of neurons, in turn, dynamically adjust to the prevailing properties of sensory stimulation, a process known as "neuronal adaptation." Neuronal adaptation is a ubiquitous phenomenon across all sensory modalities and occurs at different stages of processing from periphery to cortex. In spite of the wealth of research on contextual modulation and neuronal adaptation in visual and auditory systems, the neuronal and computational basis of sensory adaptation in somatosensory system is less understood. Here, we summarise the recent finding and views about the neuronal adaptation in the rodent whisker-mediated tactile system and further summarise the functional effect of neuronal adaptation on the response dynamics and encoding efficiency of neurons at single cell and population levels along the whisker-mediated touch system in rodents. Based on direct and indirect pieces of evidence presented here, we suggest sensory adaptation provides context-dependent functional mechanisms for noise reduction in sensory processing, salience processing and deviant stimulus detection, shift between integration and coincidence detection, band-pass frequency filtering, adjusting neuronal receptive fields, enhancing neural coding and improving discriminability around adapting stimuli, energy conservation, and disambiguating encoding of principal features of tactile stimuli.
Keywords: information theory; neural coding; neural network; neuronal adaptation; noise correlation; rodent; somatosensory; whisker system.
Copyright © 2021 Adibi and Lampl.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Neural Coding of Whisker-Mediated Touch in Primary Somatosensory Cortex Is Altered Following Early Blindness.J Neurosci. 2018 Jul 4;38(27):6172-6189. doi: 10.1523/JNEUROSCI.0066-18.2018. Epub 2018 May 28. J Neurosci. 2018. PMID: 29807911 Free PMC article.
-
Representation of tactile scenes in the rodent barrel cortex.Neuroscience. 2018 Jan 1;368:81-94. doi: 10.1016/j.neuroscience.2017.08.039. Epub 2017 Aug 23. Neuroscience. 2018. PMID: 28843997 Review.
-
Millisecond precision temporal encoding of stimulus features during cortically generated gamma oscillations in the rat somatosensory cortex.J Physiol. 2018 Feb 1;596(3):515-534. doi: 10.1113/JP275245. Epub 2018 Jan 9. J Physiol. 2018. PMID: 29265375 Free PMC article.
-
Tactile Defensiveness and Impaired Adaptation of Neuronal Activity in the Fmr1 Knock-Out Mouse Model of Autism.J Neurosci. 2017 Jul 5;37(27):6475-6487. doi: 10.1523/JNEUROSCI.0651-17.2017. Epub 2017 Jun 12. J Neurosci. 2017. PMID: 28607173 Free PMC article.
-
Neuronal adaptation in the somatosensory system of rodents.Neuroscience. 2017 Feb 20;343:66-76. doi: 10.1016/j.neuroscience.2016.11.043. Epub 2016 Dec 5. Neuroscience. 2017. PMID: 27923742 Review.
Cited by
-
Reducing Merkel cell activity in the whisker follicle disrupts cortical encoding of whisker movement amplitude and velocity.IBRO Neurosci Rep. 2022 Sep 29;13:356-363. doi: 10.1016/j.ibneur.2022.09.008. eCollection 2022 Dec. IBRO Neurosci Rep. 2022. PMID: 36281438 Free PMC article.
-
Neural mechanisms for the localization of unexpected external motion.Nat Commun. 2023 Sep 30;14(1):6112. doi: 10.1038/s41467-023-41755-z. Nat Commun. 2023. PMID: 37777516 Free PMC article.
-
Correlation of zero echo time functional MRI with neuronal activity in rats.J Cereb Blood Flow Metab. 2025 May;45(5):855-870. doi: 10.1177/0271678X251314682. Epub 2025 Jan 23. J Cereb Blood Flow Metab. 2025. PMID: 39846159 Free PMC article.
-
Whisker stimulation with different frequencies reveals non-uniform modulation of functional magnetic resonance imaging signal across sensory systems in awake rats.bioRxiv [Preprint]. 2024 Nov 14:2024.11.13.623361. doi: 10.1101/2024.11.13.623361. bioRxiv. 2024. Update in: Cereb Cortex. 2025 Jul 1;35(7):bhaf194. doi: 10.1093/cercor/bhaf194. PMID: 39605361 Free PMC article. Updated. Preprint.
-
A power law describes the magnitude of adaptation in neural populations of primary visual cortex.Nat Commun. 2023 Dec 15;14(1):8366. doi: 10.1038/s41467-023-43572-w. Nat Commun. 2023. PMID: 38102113 Free PMC article.
References
-
- Abeles M. (1982). Role of the cortical neuron: integrator or coincidence detector? Israel J. Med. Sci. 18, 83–92. - PubMed
Publication types
LinkOut - more resources
Full Text Sources