

Angiogenesis is Inhibited by Arsenic Trioxide Through Downregulation of the CircHIPK3/miR-149-5p/FOXO1/VEGF Functional Module in Rheumatoid Arthritis
- PMID: 34776969
- PMCID: PMC8579003
- DOI: 10.3389/fphar.2021.751667
Angiogenesis is Inhibited by Arsenic Trioxide Through Downregulation of the CircHIPK3/miR-149-5p/FOXO1/VEGF Functional Module in Rheumatoid Arthritis
Abstract
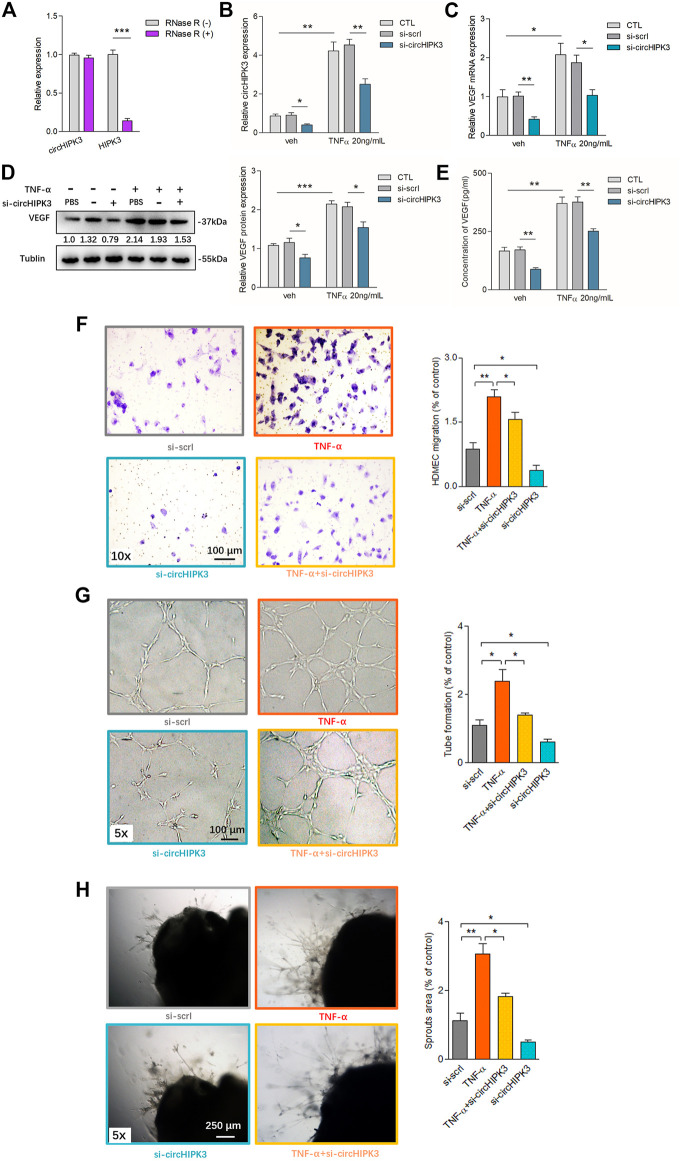
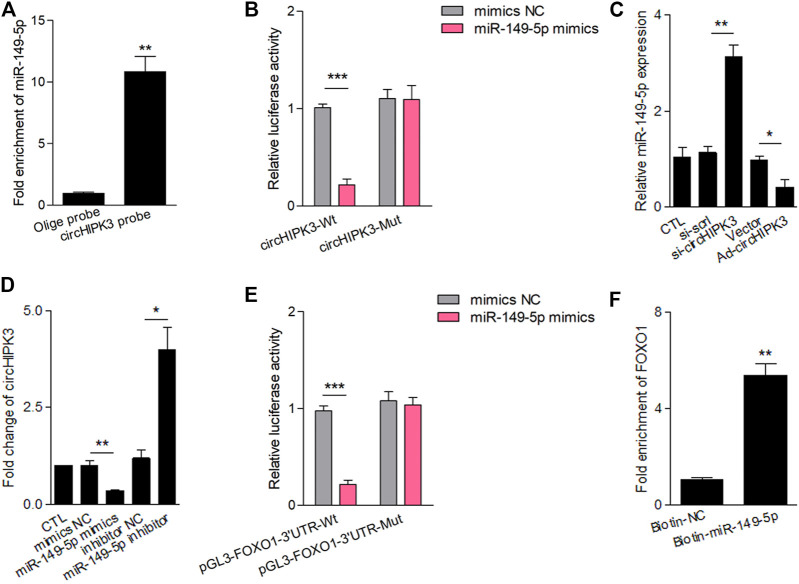
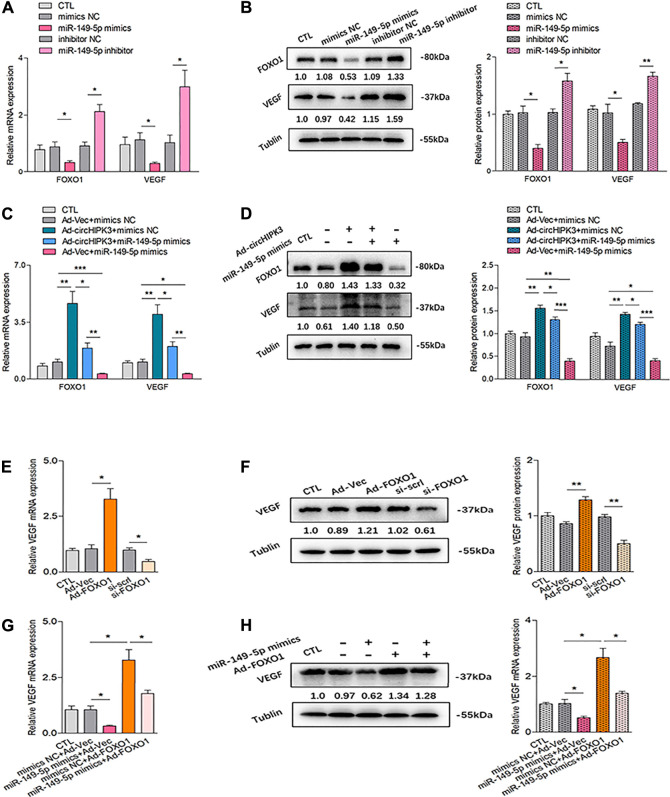
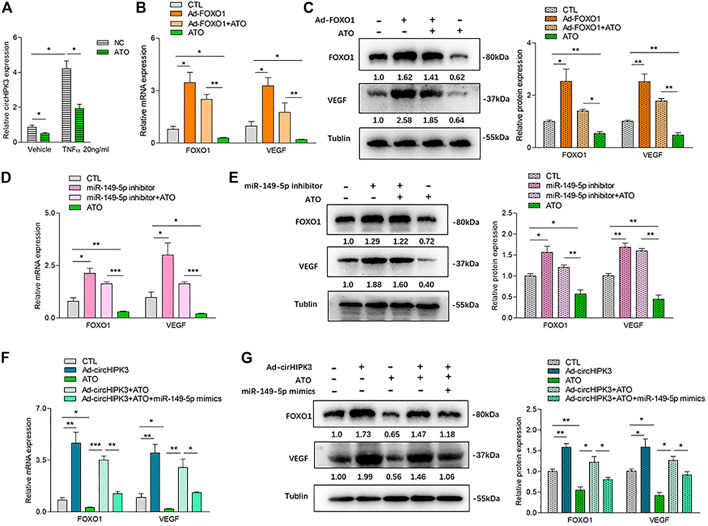
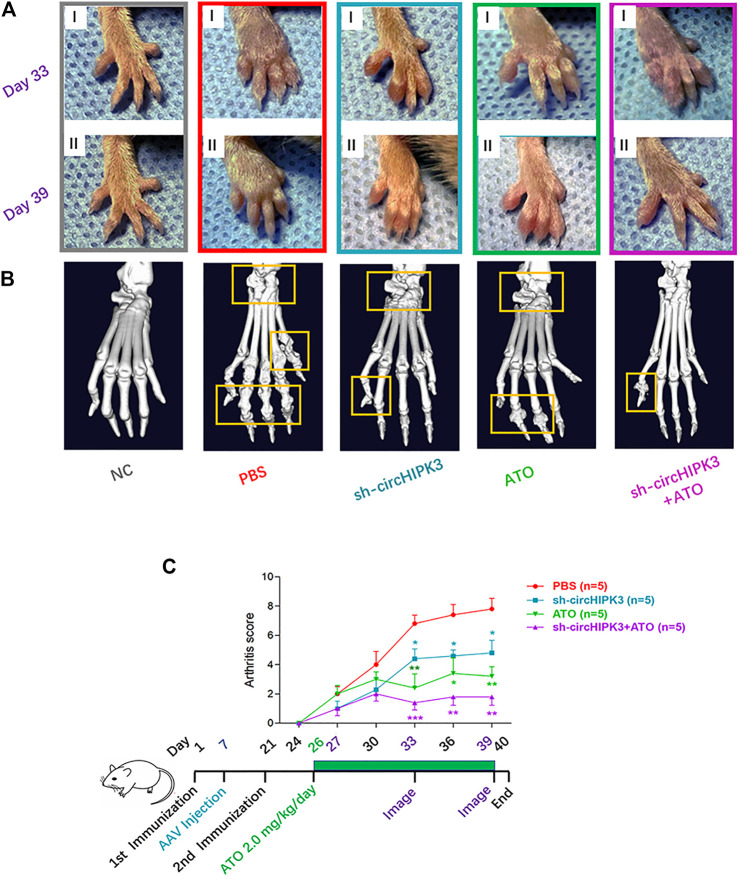
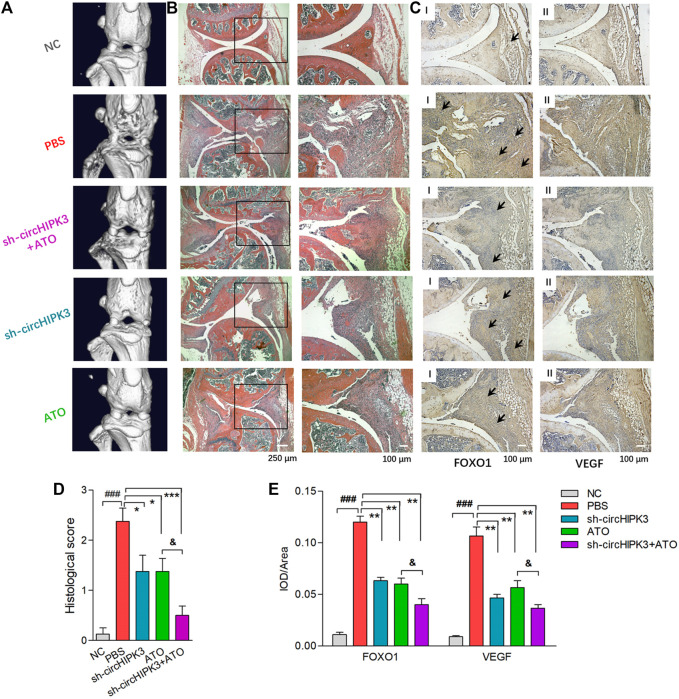
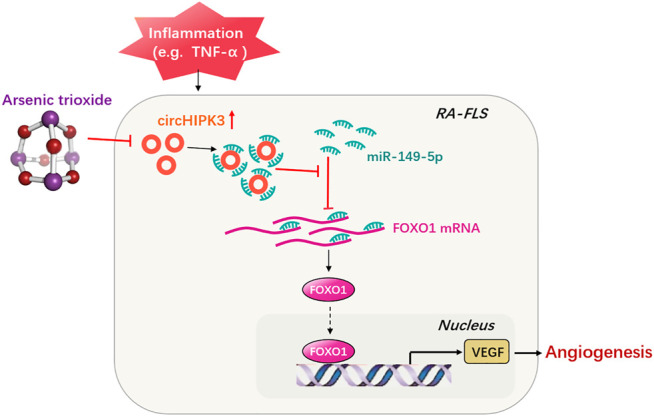
Angiogenesis is a crucial event in the pathogenesis of rheumatoid arthritis (RA). Arsenic trioxide (ATO, As2O3) has been reported to inhibit synovial angiogenesis via the vascular endothelial growth factor (VEGF)-centered functional module. However, the exact mechanisms of ATO on VEGF modulation remain unclear. Circular RNAs (circRNAs) are emerging as important regulators in RA, and the detailed mechanisms remain largely unknown. Here, we reported a circRNA (circHIPK3), the expression of which was significantly increased in RA fibroblast-like synoviocytes (RA-FLS) after TNF-α induction. Moreover, VEGF content in the supernatants of a RA-FLS and human dermal microvascular endothelial cell (HDMEC) co-culture as well as in RA-FLS co-cultured was significantly elevated in accordance with circHIPK3 levels. This increased VEGF expression may significantly upregulate endothelial tube formation and transwell migration, as well as microvessel sprouting in the ex vivo aortic ring assay. CircHIPK3 was further illustrated to be a sponge for the forkhead box transcription factor O1 (FOXO1)-targeting miR-149-5p, leading to the changing expression of the downstream VEGF. These networked factors mainly form a functional module regulating angiogenesis in RA-FLS, and the expression of this functional module could be significantly downregulated by ATO with a consistently reduced vascularity in vitro. In the collagen-induced arthritis (CIA) mice model, an intra-articular injection of the adeno-associated virus-si-circHIPK3 or ATO was demonstrated to alleviate the synovial VEGF expression and arthritis severity respectively. Thus, we elucidate a previously unknown mechanism between circRNAs and RA, and ATO has a significant protective effect on RA-FLS and CIA synovium via its inhibition of the angiogenic functional module of circHIPK3/miR-149-5p/FOXO1/VEGF, suggesting great potential for the combination therapy of ATO with circHIPK3 silencing.
Keywords: FoxO1; VEGF; angiogenesis; circHIPK3; inflammation; rheumatoid arthritis.
Copyright © 2021 Zhang, Ma, Zhang, Niu, Chu and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Dai X., Chen C., Yang Q., Xue J., Chen X., Sun B., et al. (2018). Exosomal circRNA_100284 From Arsenite-Transformed Cells, via microRNA-217 Regulation of EZH2, Is Involved in the Malignant Transformation of Human Hepatic Cells by Accelerating the Cell Cycle and Promoting Cell Proliferation. Cell Death Dis. 9 (5), 454. 10.1038/s41419-018-0485-1 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous