

Targeted protein oxidation using a chromophore-modified rapamycin analog
- PMID: 34777761
- PMCID: PMC8528027
- DOI: 10.1039/d1sc04464h
Targeted protein oxidation using a chromophore-modified rapamycin analog
Abstract
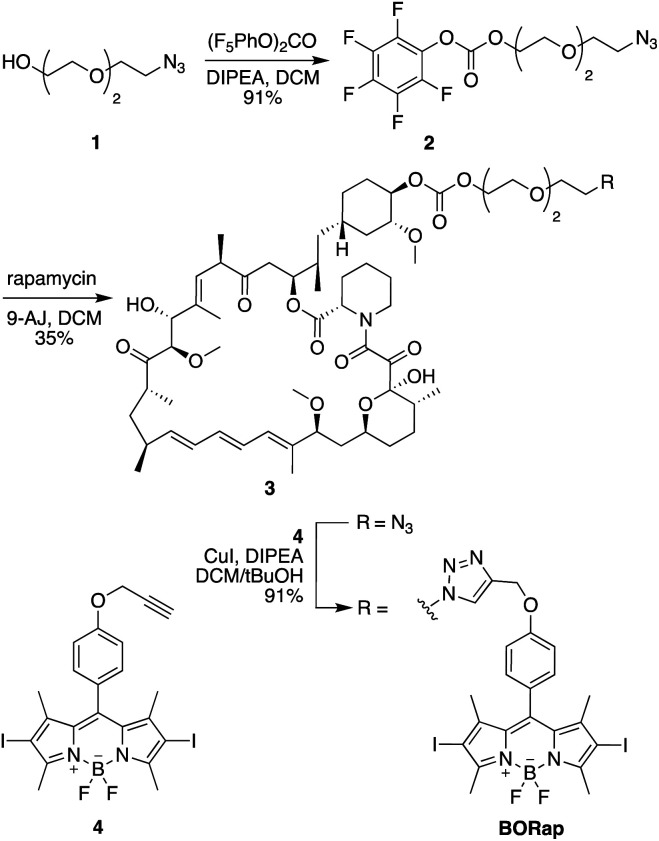
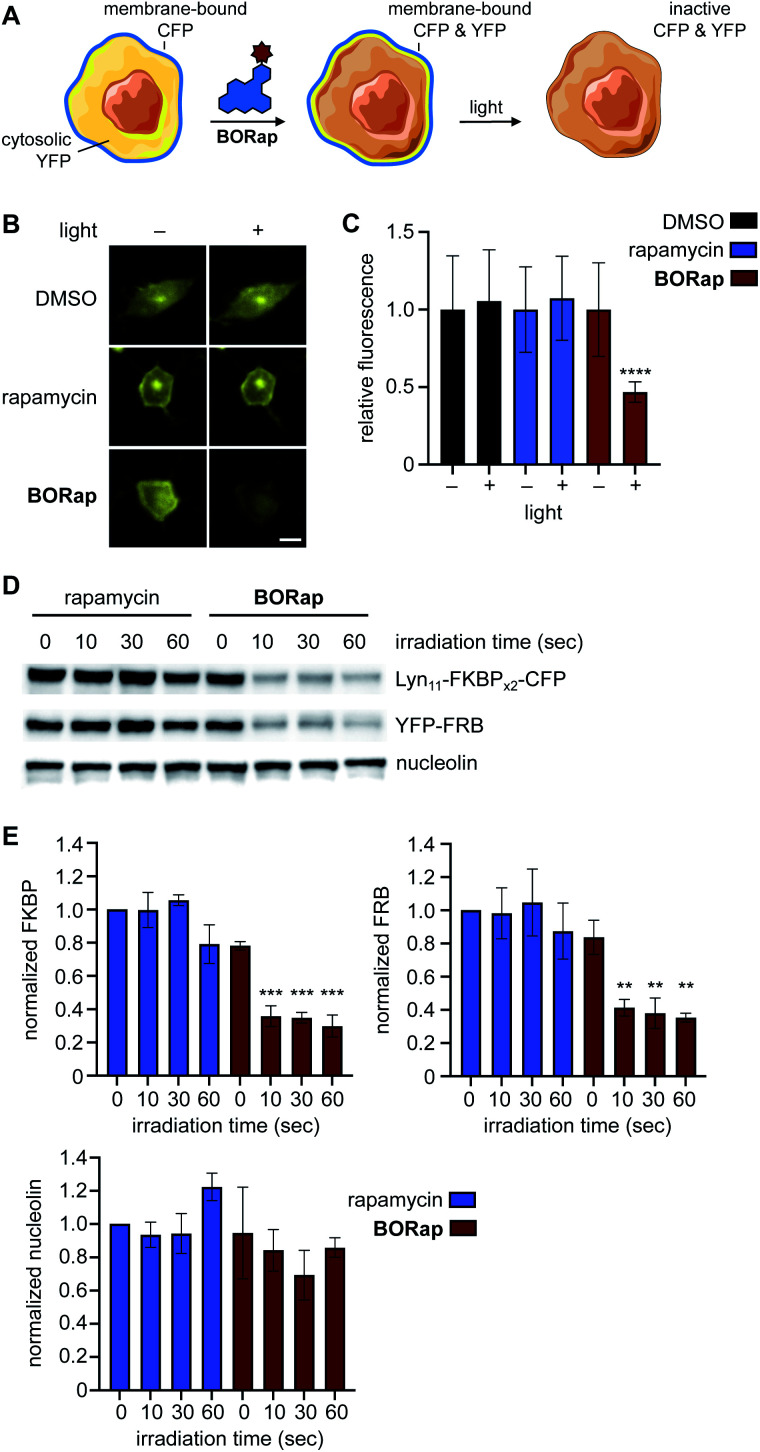
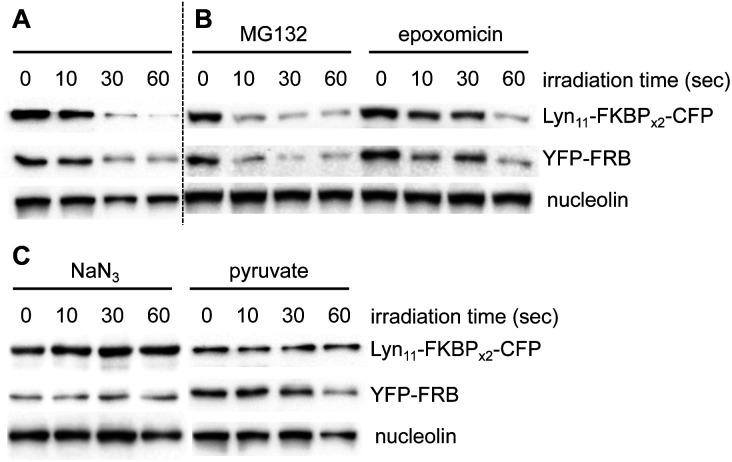
Chemically induced dimerization of FKBP and FRB using rapamycin and rapamycin analogs has been utilized in a variety of biological applications. Formation of the FKBP-rapamycin-FRB ternary complex is typically used to activate a biological process and this interaction has proven to be essentially irreversible. In many cases, it would be beneficial to also have temporal control over deactivating a biological process once it has been initiated. Thus, we developed the first reactive oxygen species-generating rapamycin analog toward this goal. The BODIPY-rapamycin analog BORap is capable of dimerizing FKBP and FRB to form a ternary complex, and upon irradiation with 530 nm light, generates singlet oxygen to oxidize and inactivate proteins of interest fused to FKBP/FRB.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts of interest to declare.
Figures


Similar articles
-
Synthesis and application of light-switchable arylazopyrazole rapamycin analogs.Org Biomol Chem. 2019 Sep 28;17(36):8348-8353. doi: 10.1039/c9ob01719d. Epub 2019 Aug 30. Org Biomol Chem. 2019. PMID: 31469140
-
Characterization of the FKBP.rapamycin.FRB ternary complex.J Am Chem Soc. 2005 Apr 6;127(13):4715-21. doi: 10.1021/ja043277y. J Am Chem Soc. 2005. PMID: 15796538
-
Structural insights into rapamycin-induced oligomerization of a FRB-FKBP fusion protein.FEBS Lett. 2024 Sep;598(18):2292-2305. doi: 10.1002/1873-3468.14986. Epub 2024 Jul 19. FEBS Lett. 2024. PMID: 39031920
-
Applications of genetically encoded photosensitizer miniSOG: from correlative light electron microscopy to immunophotosensitizing.J Biophotonics. 2017 Mar;10(3):338-352. doi: 10.1002/jbio.201600120. Epub 2016 Jul 20. J Biophotonics. 2017. PMID: 27435584 Review.
-
Synthesis and biological evaluation of rapamycin-derived, next generation small molecules.Medchemcomm. 2017 Nov 22;9(1):27-43. doi: 10.1039/c7md00474e. eCollection 2018 Jan 1. Medchemcomm. 2017. PMID: 30108899 Free PMC article. Review.
References
-
- Wang P. Gao Z. Yuan M. Zhu J. Wang F. Mechanically linked poly[2]rotaxanes constructed from the benzo-21-crown-7/secondary ammonium salt recognition motif. Polym. Chem. 2016;7(22):3664–3668. doi: 10.1039/C6PY00494F. - DOI
LinkOut - more resources
Full Text Sources
Research Materials