

Progress in Gene Editing Tools and Their Potential for Correcting Mutations Underlying Hearing and Vision Loss
- PMID: 34778871
- PMCID: PMC8581640
- DOI: 10.3389/fgeed.2021.737632
Progress in Gene Editing Tools and Their Potential for Correcting Mutations Underlying Hearing and Vision Loss
Abstract
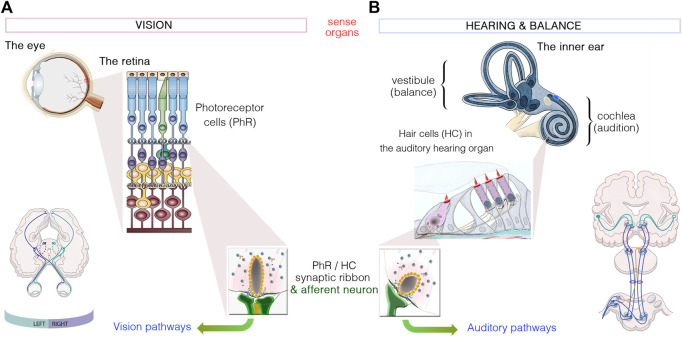
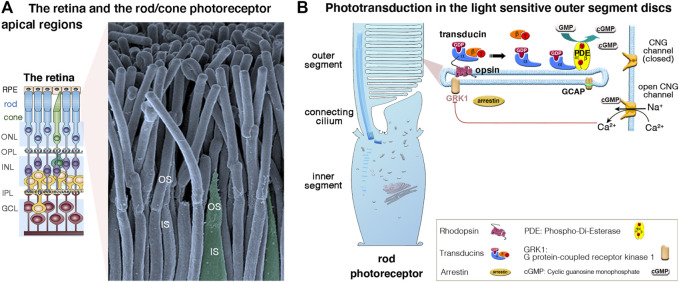
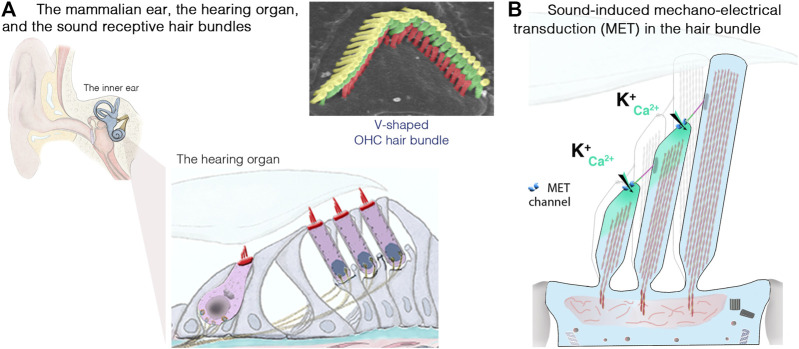
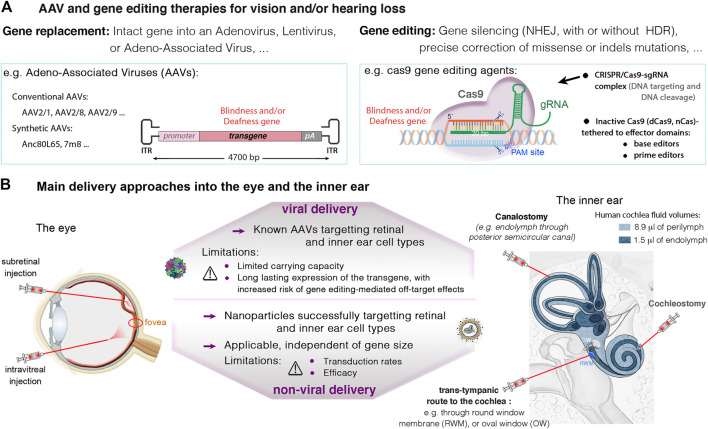
Blindness and deafness are the most frequent sensory disorders in humans. Whatever their cause - genetic, environmental, or due to toxic agents, or aging - the deterioration of these senses is often linked to irreversible damage to the light-sensing photoreceptor cells (blindness) and/or the mechanosensitive hair cells (deafness). Efforts are increasingly focused on preventing disease progression by correcting or replacing the blindness and deafness-causal pathogenic alleles. In recent years, gene replacement therapies for rare monogenic disorders of the retina have given positive results, leading to the marketing of the first gene therapy product for a form of childhood hereditary blindness. Promising results, with a partial restoration of auditory function, have also been reported in preclinical models of human deafness. Silencing approaches, including antisense oligonucleotides, adeno-associated virus (AAV)-mediated microRNA delivery, and genome-editing approaches have also been applied to various genetic forms of blindness and deafness The discovery of new DNA- and RNA-based CRISPR/Cas nucleases, and the new generations of base, prime, and RNA editors offers new possibilities for directly repairing point mutations and therapeutically restoring gene function. Thanks to easy access and immune-privilege status of self-contained compartments, the eye and the ear continue to be at the forefront of developing therapies for genetic diseases. Here, we review the ongoing applications and achievements of this new class of emerging therapeutics in the sensory organs of vision and hearing, highlighting the challenges ahead and the solutions to be overcome for their successful therapeutic application in vivo.
Keywords: CRISPR/Cas9; blindness; deafness (hearing loss); gene editing; gene therapy; hair cells; inherited retinal degeneration (IRD); retina.
Copyright © 2021 Botto, Dalkara and El-Amraoui.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The Opportunities and Challenges of Gene Therapy for Treatment of Inherited Forms of Vision and Hearing Loss.Hum Gene Ther. 2023 Sep;34(17-18):808-820. doi: 10.1089/hum.2023.126. Hum Gene Ther. 2023. PMID: 37642267 Review.
-
Inner Ear Gene Therapies Take Off: Current Promises and Future Challenges.J Clin Med. 2020 Jul 21;9(7):2309. doi: 10.3390/jcm9072309. J Clin Med. 2020. PMID: 32708116 Free PMC article. Review.
-
CRISPR/Cas9: targeted genome editing for the treatment of hereditary hearing loss.J Appl Genet. 2020 Feb;61(1):51-65. doi: 10.1007/s13353-019-00535-6. Epub 2020 Jan 7. J Appl Genet. 2020. PMID: 31912450 Review.
-
In vivo base editing restores sensory transduction and transiently improves auditory function in a mouse model of recessive deafness.Sci Transl Med. 2020 Jun 3;12(546):eaay9101. doi: 10.1126/scitranslmed.aay9101. Sci Transl Med. 2020. PMID: 32493795 Free PMC article.
-
Advances in gene therapy hold promise for treating hereditary hearing loss.Mol Ther. 2023 Apr 5;31(4):934-950. doi: 10.1016/j.ymthe.2023.02.001. Epub 2023 Feb 8. Mol Ther. 2023. PMID: 36755494 Free PMC article. Review.
Cited by
-
Gene Therapy for Rhodopsin Mutations.Cold Spring Harb Perspect Med. 2022 Aug 8;12(9):a041283. doi: 10.1101/cshperspect.a041283. Online ahead of print. Cold Spring Harb Perspect Med. 2022. PMID: 35940643 Free PMC article.
-
G protein-coupled receptors in cochlea: Potential therapeutic targets for hearing loss.Front Mol Neurosci. 2022 Oct 12;15:1028125. doi: 10.3389/fnmol.2022.1028125. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36311029 Free PMC article. Review.
-
Promising Strategies for Preserving Adult Endothelium Health and Reversing Its Dysfunction: From Liquid Biopsy to New Omics Technologies and Noninvasive Circulating Biomarkers.Int J Mol Sci. 2022 Jul 7;23(14):7548. doi: 10.3390/ijms23147548. Int J Mol Sci. 2022. PMID: 35886888 Free PMC article. Review.
-
Poly (β-amino Ester) Nanoparticles Modified with a Rabies Virus-derived peptide for the Delivery of ASCL1 Across a 3D In Vitro Model of the Blood Brain Barrier.ACS Appl Nano Mater. 2023 Apr 14;6(7):6299-6311. doi: 10.1021/acsanm.3c00651. Epub 2023 Mar 31. ACS Appl Nano Mater. 2023. PMID: 37274933 Free PMC article.
-
Dual AAV-based PCDH15 gene therapy achieves sustained rescue of visual function in a mouse model of Usher syndrome 1F.Mol Ther. 2023 Dec 6;31(12):3490-3501. doi: 10.1016/j.ymthe.2023.10.017. Epub 2023 Oct 20. Mol Ther. 2023. PMID: 37864333 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources