

The functional characterization of callosal connections
- PMID: 34780864
- PMCID: PMC8752969
- DOI: 10.1016/j.pneurobio.2021.102186
The functional characterization of callosal connections
Abstract
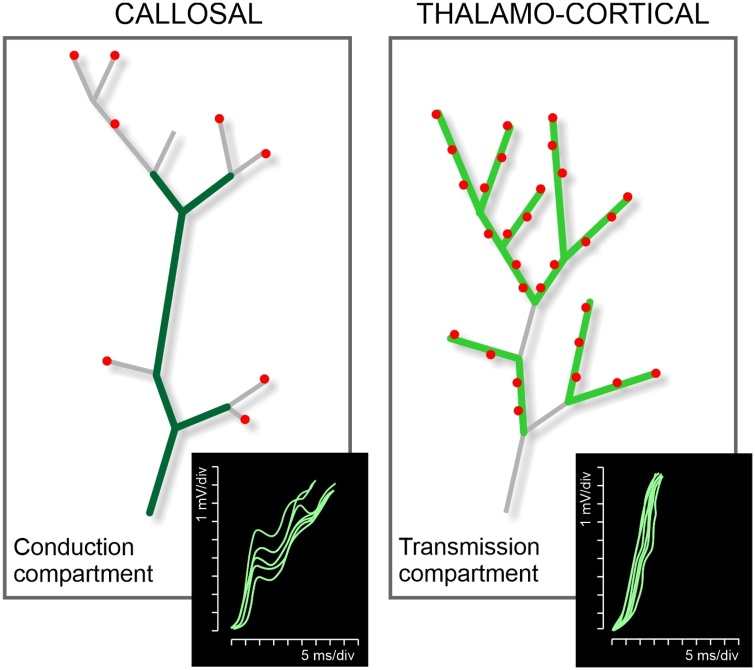
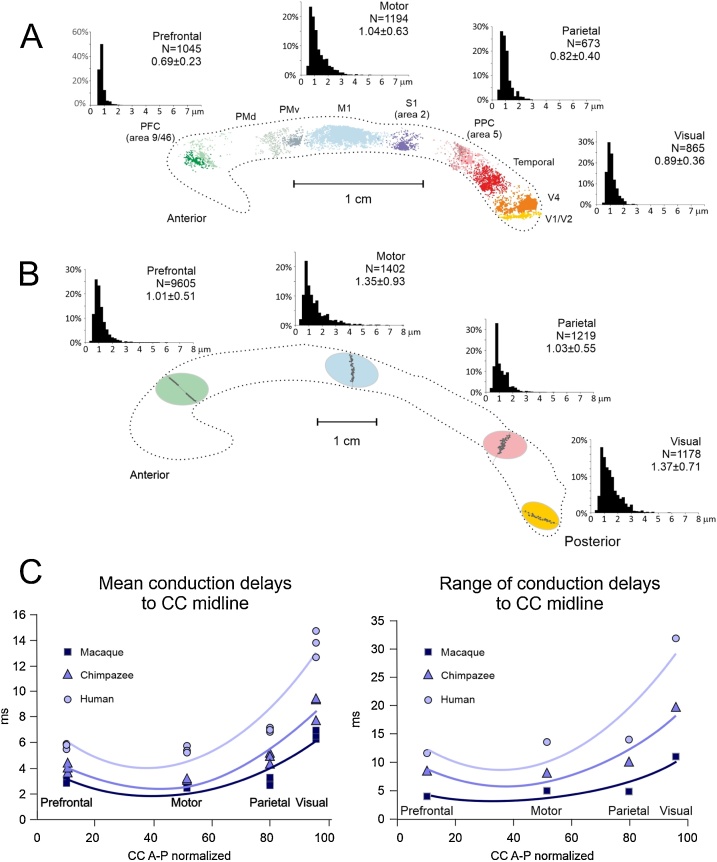
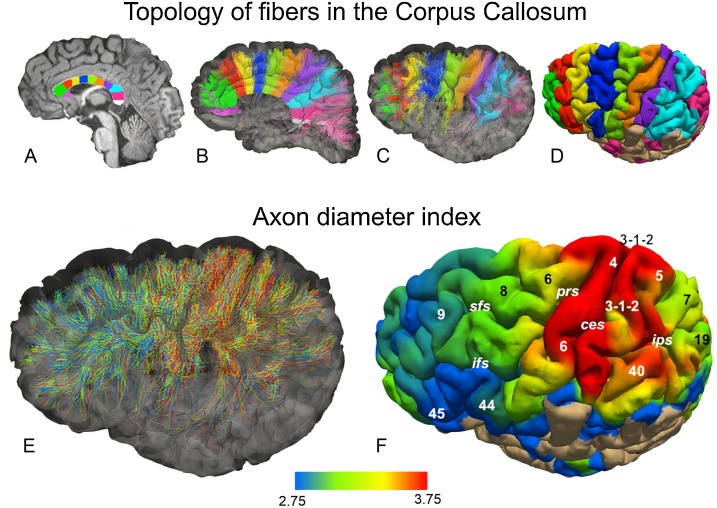
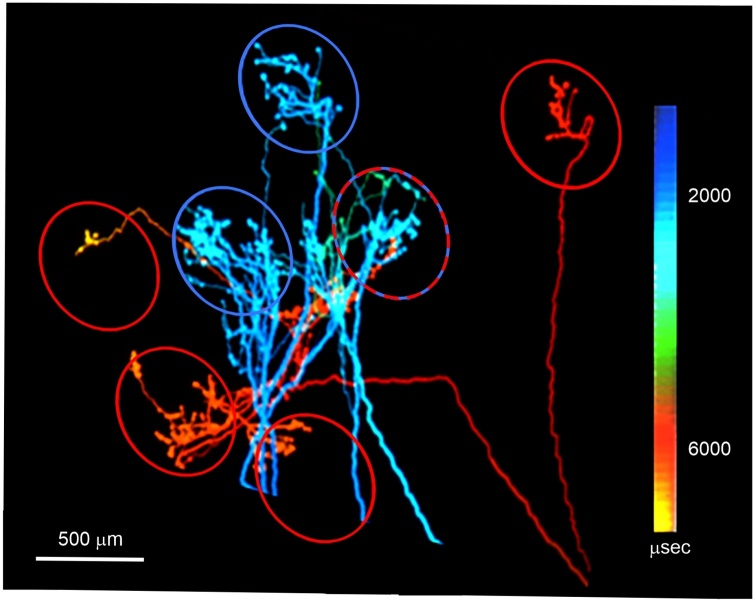
The brain operates through the synaptic interaction of distant neurons within flexible, often heterogeneous, distributed systems. Histological studies have detailed the connections between distant neurons, but their functional characterization deserves further exploration. Studies performed on the corpus callosum in animals and humans are unique in that they capitalize on results obtained from several neuroscience disciplines. Such data inspire a new interpretation of the function of callosal connections and delineate a novel road map, thus paving the way toward a general theory of cortico-cortical connectivity. Here we suggest that callosal axons can drive their post-synaptic targets preferentially when coupled to other inputs endowing the cortical network with a high degree of conditionality. This might depend on several factors, such as their pattern of convergence-divergence, the excitatory and inhibitory operation mode, the range of conduction velocities, the variety of homotopic and heterotopic projections and, finally, the state-dependency of their firing. We propose that, in addition to direct stimulation of post-synaptic targets, callosal axons often play a conditional driving or modulatory role, which depends on task contingencies, as documented by several recent studies.
Keywords: Callosal axon diameter; Callosal conduction velocity; Callosal connections flexibility; Callosal disconnection syndromes; Callosal interhemispheric transfer; Corpus callosum.
Copyright © 2021 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests
Figures
References
-
- Abeles M. Role of the cortical neuron: integrator or coincidence detector? Isr. J. Med. Sci. 1982;18:83–92. - PubMed
-
- Aboitiz F., Scheibel A.B., Fisher R.S., Zaidel E. Fiber composition of the human corpus callosum. Brain Res. 1992;598:143–153. - PubMed
-
- Akelaitis A. Studies on the corpus callosum. II. The higher visual functions in each homonymous field following complete section of the corpus callosum. Arch. Neurol. Psychiatry. 1941;45:788–796. doi: 10.1001/archneurpsyc.1941.02280170066005. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
