

HIV Proviral Burden, Genetic Diversity, and Dynamics in Viremic Controllers Who Subsequently Initiated Suppressive Antiretroviral Therapy
- PMID: 34781741
- PMCID: PMC8693448
- DOI: 10.1128/mBio.02490-21
HIV Proviral Burden, Genetic Diversity, and Dynamics in Viremic Controllers Who Subsequently Initiated Suppressive Antiretroviral Therapy
Abstract
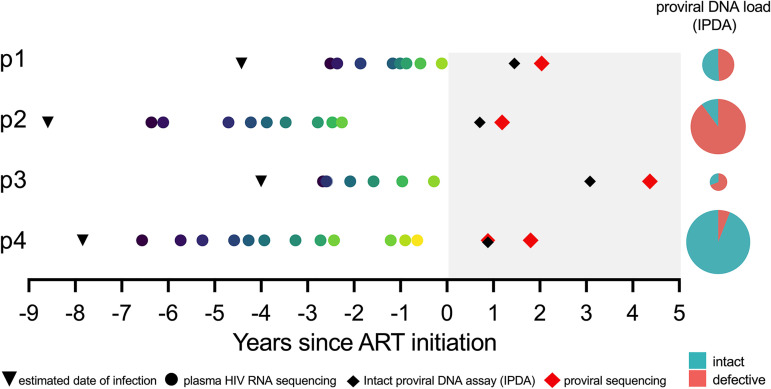
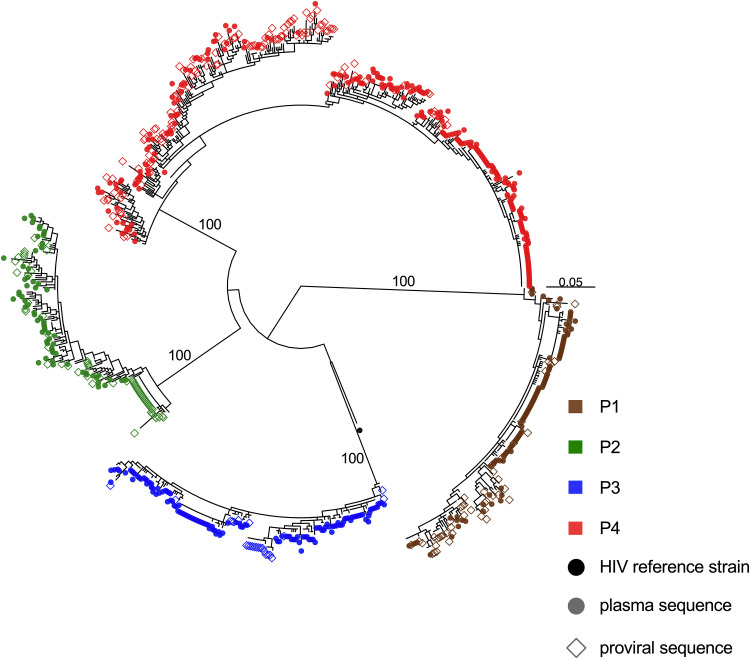
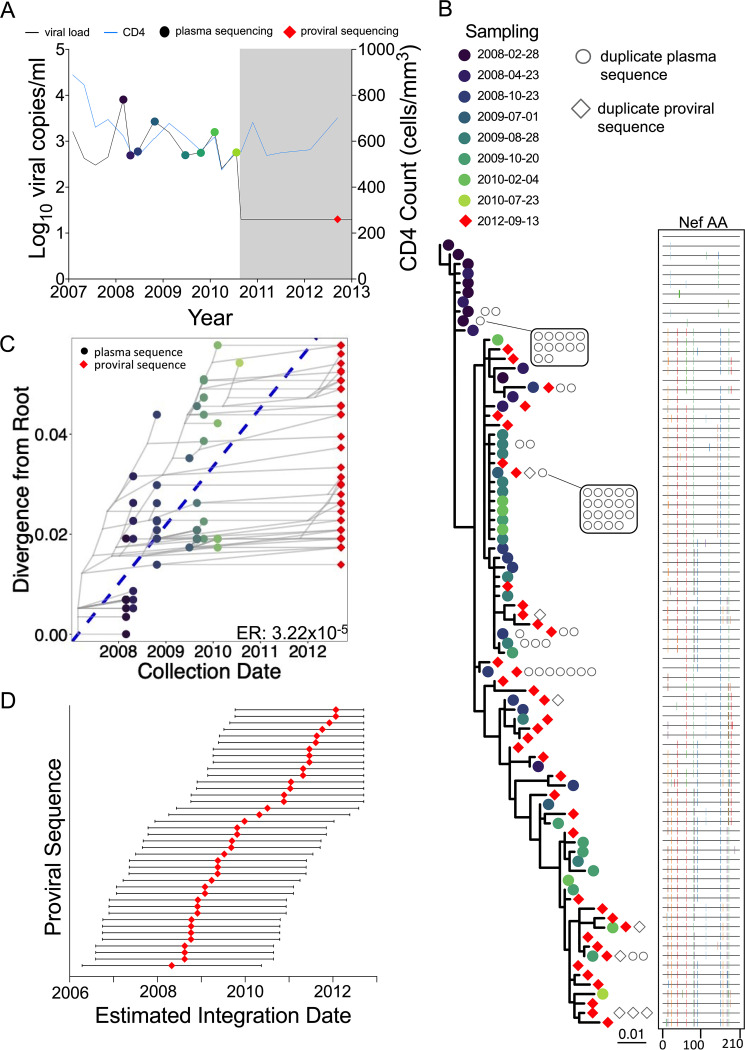
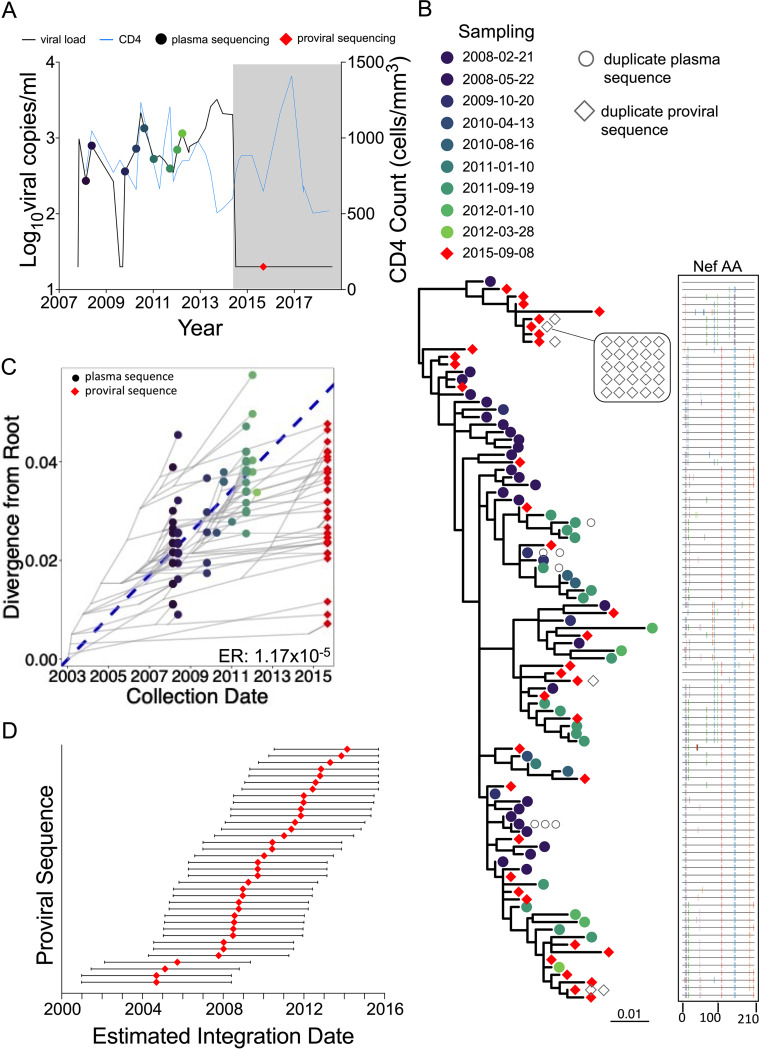
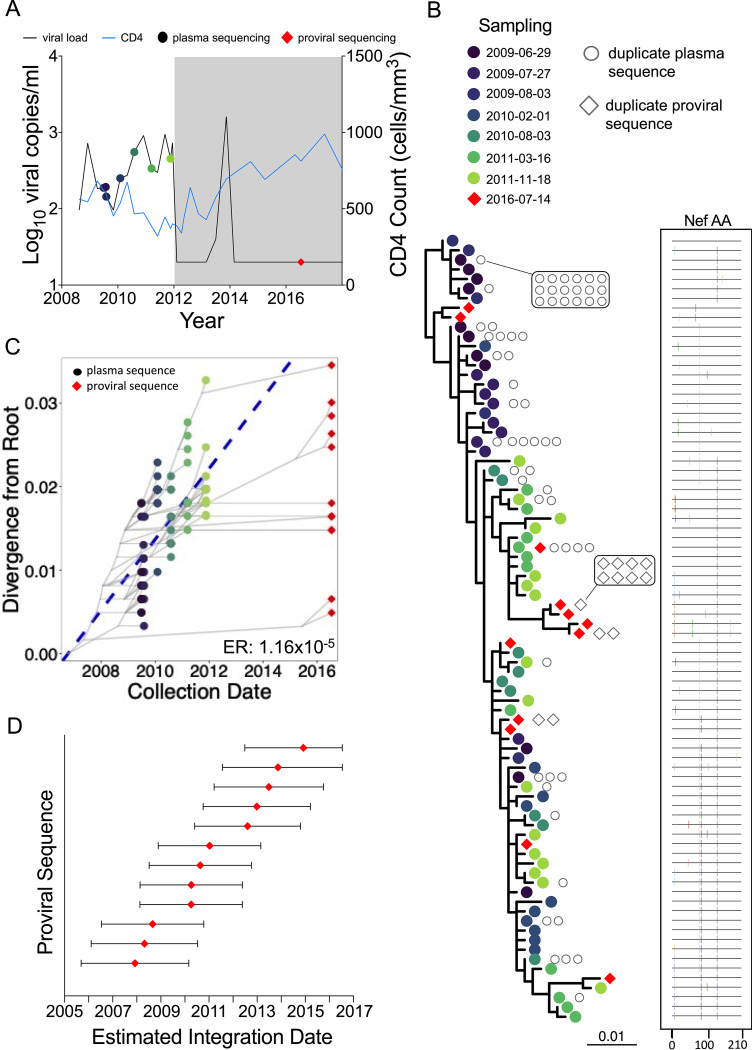
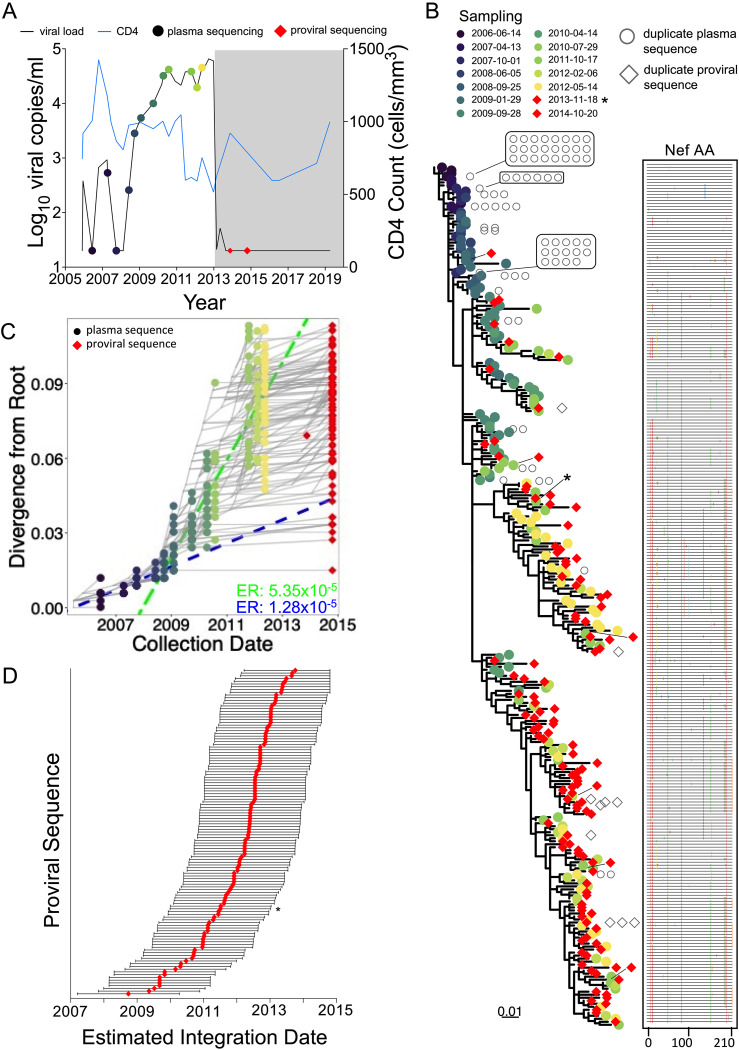
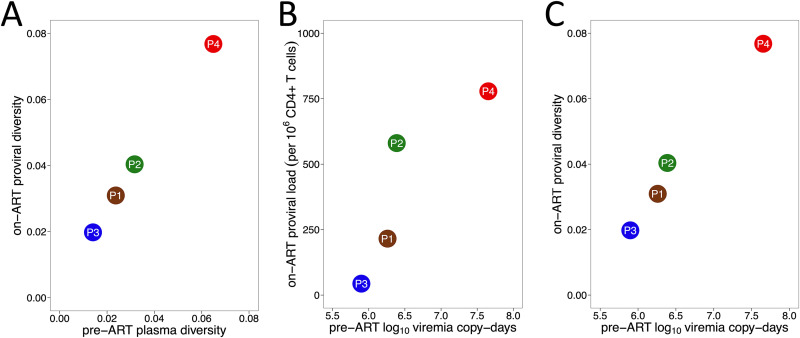
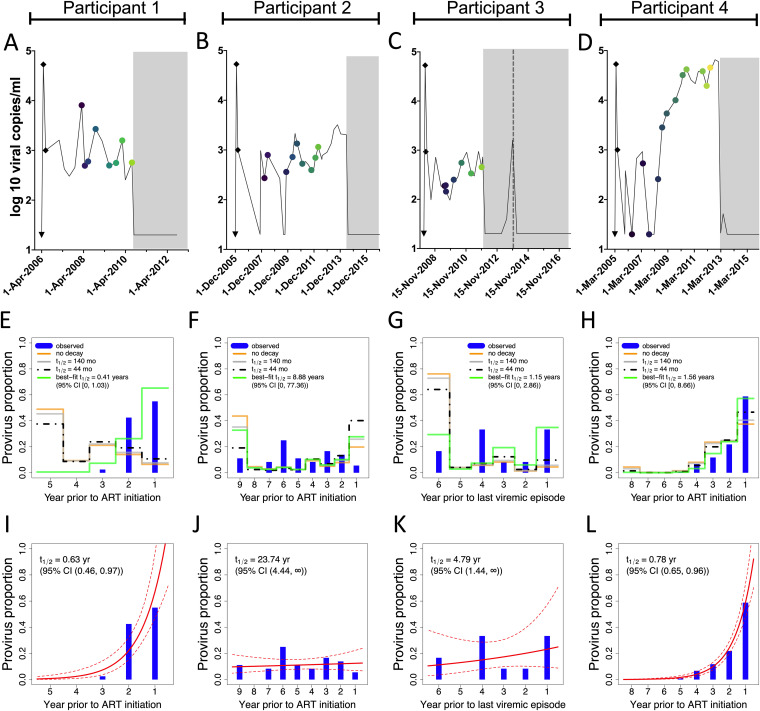
Curing HIV will require eliminating the reservoir of integrated, replication-competent proviruses that persist despite antiretroviral therapy (ART). Understanding the burden, genetic diversity, and longevity of persisting proviruses in diverse individuals with HIV is critical to this goal, but these characteristics remain understudied in some groups. Among them are viremic controllers-individuals who naturally suppress HIV to low levels but for whom therapy is nevertheless recommended. We reconstructed within-host HIV evolutionary histories from longitudinal single-genome amplified viral sequences in four viremic controllers who eventually initiated ART and used this information to characterize the age and diversity of proviruses persisting on therapy. We further leveraged these within-host proviral age distributions to estimate rates of proviral turnover prior to ART. This is an important yet understudied metric, since pre-ART proviral turnover dictates reservoir composition at ART initiation (and thereafter), which is when curative interventions, once developed, would be administered. Despite natural viremic control, all participants displayed significant within-host HIV evolution pretherapy, where overall on-ART proviral burden and diversity broadly reflected the extent of viral replication and diversity pre-ART. Consistent with recent studies of noncontrollers, the proviral pools of two participants were skewed toward sequences that integrated near ART initiation, suggesting dynamic proviral turnover during untreated infection. In contrast, proviruses recovered from the other two participants dated to time points that were more evenly spread throughout infection, suggesting slow or negligible proviral decay following deposition. HIV cure strategies will need to overcome within-host proviral diversity, even in individuals who naturally controlled HIV replication before therapy. IMPORTANCE HIV therapy is lifelong because integrated, replication-competent viral copies persist within long-lived cells. To cure HIV, we need to understand when these viral reservoirs form, how large and genetically diverse they are, and how long they endure. Elite controllers-individuals who naturally suppress HIV to undetectable levels-are being intensely studied as models of HIV remission, but viremic controllers, individuals who naturally suppress HIV to low levels, remain understudied even though they too may hold valuable insights. We combined phylogenetics and mathematical modeling to reconstruct proviral seeding and decay from infection to therapy-mediated suppression in four viremic controllers. We recovered diverse proviruses persisting during therapy that broadly reflected HIV's within-host evolutionary history, where the estimated half-lives of the persistent proviral pool during untreated infection ranged from <1 year to negligible. Cure strategies will need to contend with proviral diversity and between-host heterogeneity, even in individuals who naturally control HIV.
Keywords: genetic diversity; human immunodeficiency virus; proviral burden and dynamics; proviral half-life; viremic controllers.
Figures
References
-
- Finzi D, Hermankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, Gallant J, Markowitz M, Ho DD, Richman DD, Siliciano RF. 1997. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278:1295–1300. doi: 10.1126/science.278.5341.1295. - DOI - PubMed
-
- Bruner KM, Murray AJ, Pollack RA, Soliman MG, Laskey SB, Capoferri AA, Lai J, Strain MC, Lada SM, Hoh R, Ho YC, Richman DD, Deeks SG, Siliciano JD, Siliciano RF. 2016. Defective proviruses rapidly accumulate during acute HIV-1 infection. Nat Med 22:1043–1049. doi: 10.1038/nm.4156. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
