

Bi-allelic variants in human WDR63 cause male infertility via abnormal inner dynein arms assembly
- PMID: 34782613
- PMCID: PMC8593051
- DOI: 10.1038/s41421-021-00327-5
Bi-allelic variants in human WDR63 cause male infertility via abnormal inner dynein arms assembly
Abstract
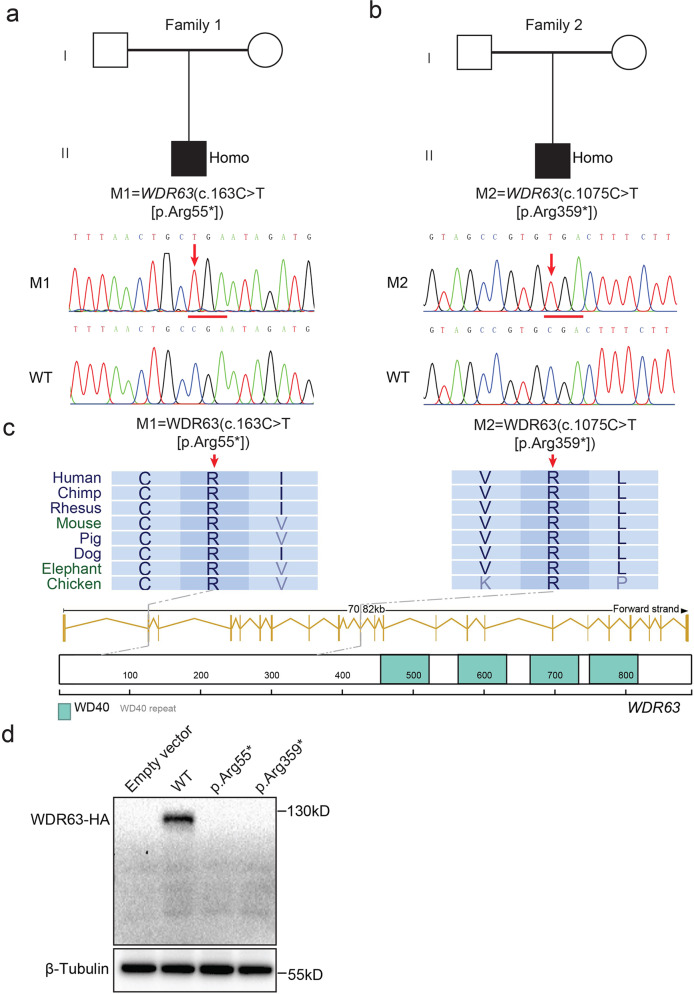
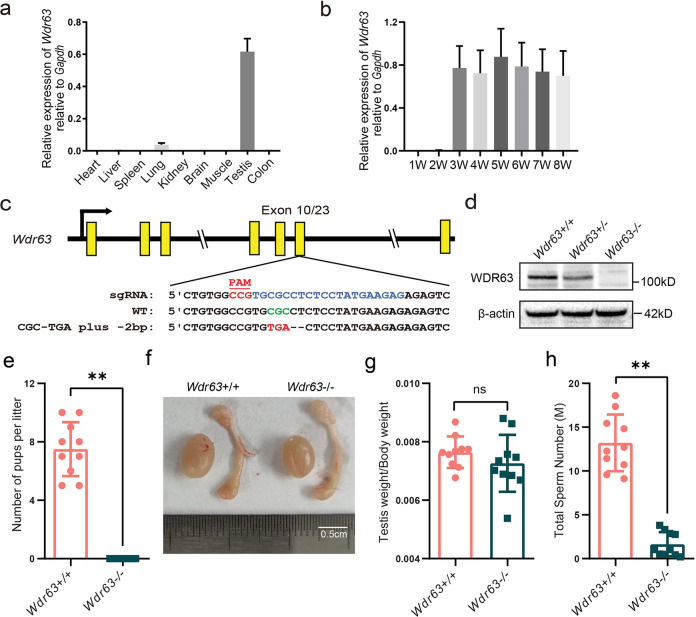
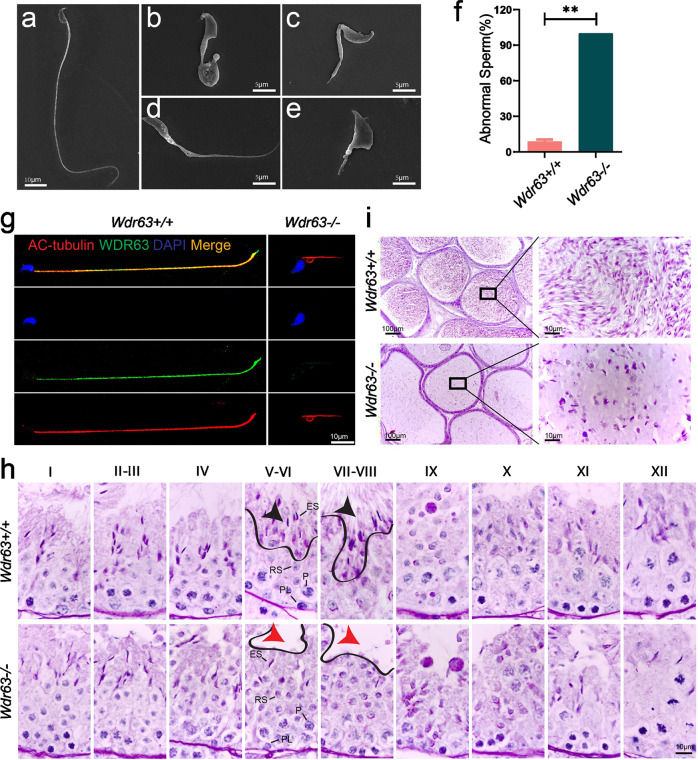
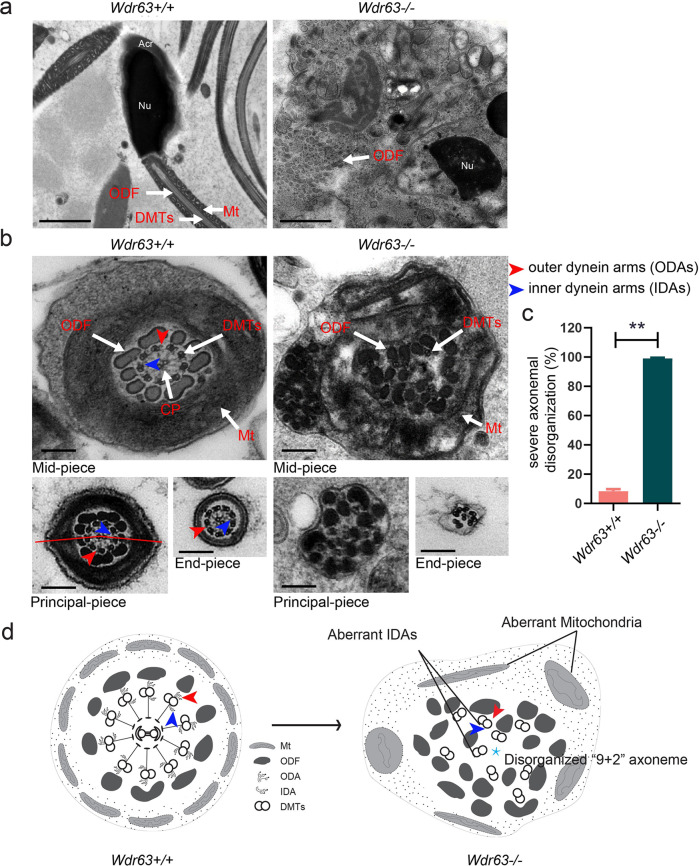
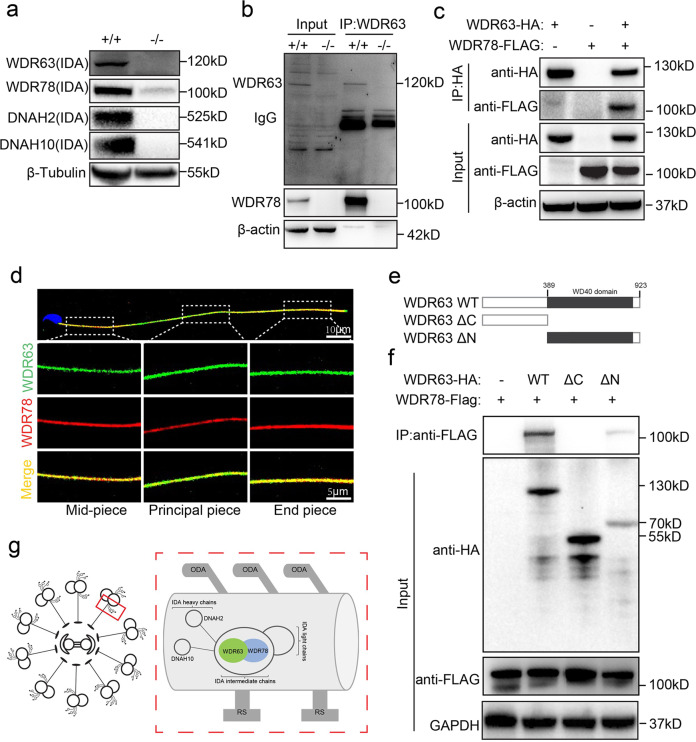
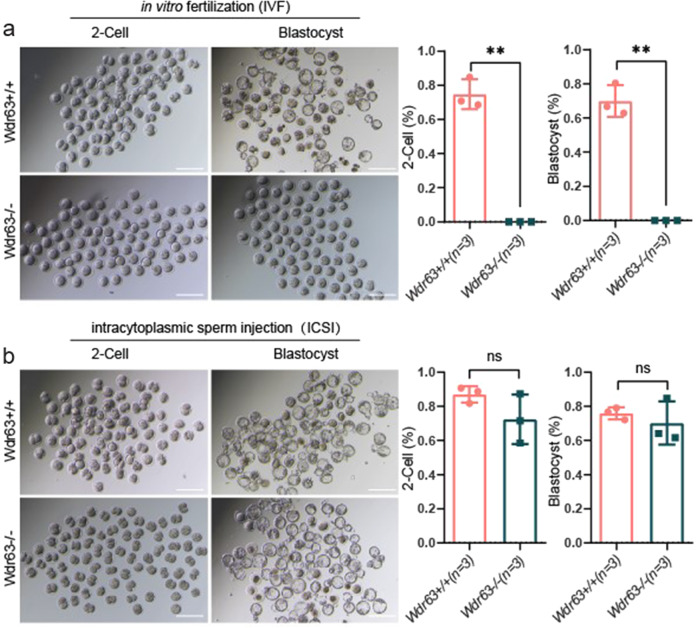
Inner dynein arm (IDA), composed of a series of protein complex, is necessary to cilia and flagella bend formation and beating. Previous studies indicated that defects of IDA protein complex result in multiple morphological abnormalities of the sperm flagellum (MMAF) and male infertility. However, the genetic causes and molecular mechanisms in the IDAs need further exploration. Here we identified two loss-of-function variants of WDR63 in both MMAF and non-obstructive azoospermia (NOA) affected cohorts. WDR63 encodes an IDA-associated protein that is dominantly expressed in testis. We next generated Wdr63-knockout (Wdr63-KO) mice through the CRISPR-Cas9 technology. Remarkably, Wdr63-KO induced decreased sperm number, abnormal flagellar morphology and male infertility. In addition, transmission electron microscopy assay showed severely disorganized "9 + 2" axoneme and absent inner dynein arms in the spermatozoa from Wdr63-KO male mice. Mechanistically, we found that WDR63 interacted with WDR78 mainly via WD40-repeat domain and is necessary for IDA assembly. Furthermore, WDR63-associated male infertility in human and mice could be overcome by intracytoplasmic sperm injection (ICSI) treatment. In conclusion, the present study demonstrates that bi-allelic variants of WDR63 cause male infertility via abnormal inner dynein arms assembly and flagella formation and can be used as a genetic diagnostic indicator for infertility males.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
TCTEX1D2 is essential for sperm flagellum formation in mice.Sci Rep. 2025 Jan 18;15(1):2413. doi: 10.1038/s41598-024-83424-1. Sci Rep. 2025. PMID: 39827215 Free PMC article.
-
Bi-allelic mutations of DNAH10 cause primary male infertility with asthenoteratozoospermia in humans and mice.Am J Hum Genet. 2021 Aug 5;108(8):1466-1477. doi: 10.1016/j.ajhg.2021.06.010. Epub 2021 Jul 7. Am J Hum Genet. 2021. PMID: 34237282 Free PMC article.
-
Defects in the cytoplasmic assembly of axonemal dynein arms cause morphological abnormalities and dysmotility in sperm cells leading to male infertility.PLoS Genet. 2021 Feb 26;17(2):e1009306. doi: 10.1371/journal.pgen.1009306. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33635866 Free PMC article.
-
Composition and function of ciliary inner-dynein-arm subunits studied in Chlamydomonas reinhardtii.Cytoskeleton (Hoboken). 2021 Mar;78(3):77-96. doi: 10.1002/cm.21662. Epub 2021 Apr 28. Cytoskeleton (Hoboken). 2021. PMID: 33876572 Free PMC article. Review.
-
Novel DNAH1 Mutation Loci Lead to Multiple Morphological Abnormalities of the Sperm Flagella and Literature Review.World J Mens Health. 2022 Oct;40(4):551-560. doi: 10.5534/wjmh.210119. Epub 2022 Jan 25. World J Mens Health. 2022. PMID: 35118838 Free PMC article. Review.
Cited by
-
Dnajc5b contributes to male fertility by maintaining the mitochondrial functions and autophagic homeostasis during spermiogenesis.Cell Mol Life Sci. 2025 Feb 3;82(1):69. doi: 10.1007/s00018-024-05552-1. Cell Mol Life Sci. 2025. PMID: 39899042 Free PMC article.
-
Identification of CFAP52 as a novel diagnostic target of male infertility with defects of sperm head-tail connection and flagella development.Elife. 2023 Dec 21;12:RP92769. doi: 10.7554/eLife.92769. Elife. 2023. PMID: 38126872 Free PMC article.
-
Homozygous CCDC146 mutation causes oligoasthenoteratozoospermia in humans and mice.Zool Res. 2024 Sep 18;45(5):1073-1087. doi: 10.24272/j.issn.2095-8137.2024.019. Zool Res. 2024. PMID: 39245651 Free PMC article.
-
Homozygous deleterious variants in MYCBPAP induce asthenoteratozoospermia involving abnormal acrosome biogenesis, manchette structure and sperm tail assembly in humans and mice.Sci China Life Sci. 2025 Mar;68(3):777-792. doi: 10.1007/s11427-024-2757-7. Epub 2024 Dec 18. Sci China Life Sci. 2025. PMID: 39704931
-
TCTEX1D2 is essential for sperm flagellum formation in mice.Sci Rep. 2025 Jan 18;15(1):2413. doi: 10.1038/s41598-024-83424-1. Sci Rep. 2025. PMID: 39827215 Free PMC article.
References
-
- Toure A, et al. The genetic architecture of morphological abnormalities of the sperm tail. Hum. Genet. 2021;140:21–42. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
