

α2δ-2 is required for depolarization-induced suppression of excitation in Purkinje cells
- PMID: 34783012
- PMCID: PMC8724408
- DOI: 10.1113/JP282438
α2δ-2 is required for depolarization-induced suppression of excitation in Purkinje cells
Abstract
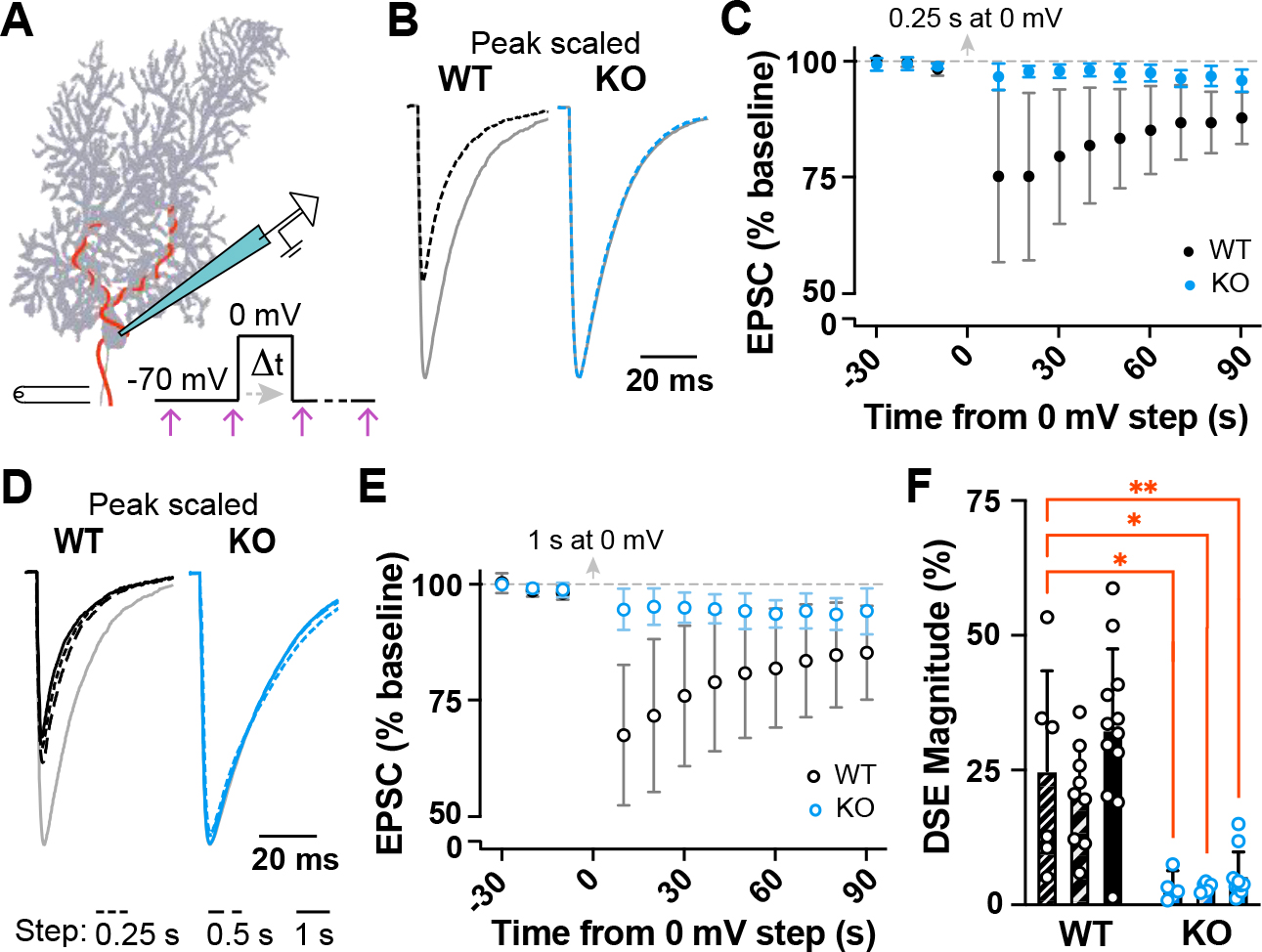
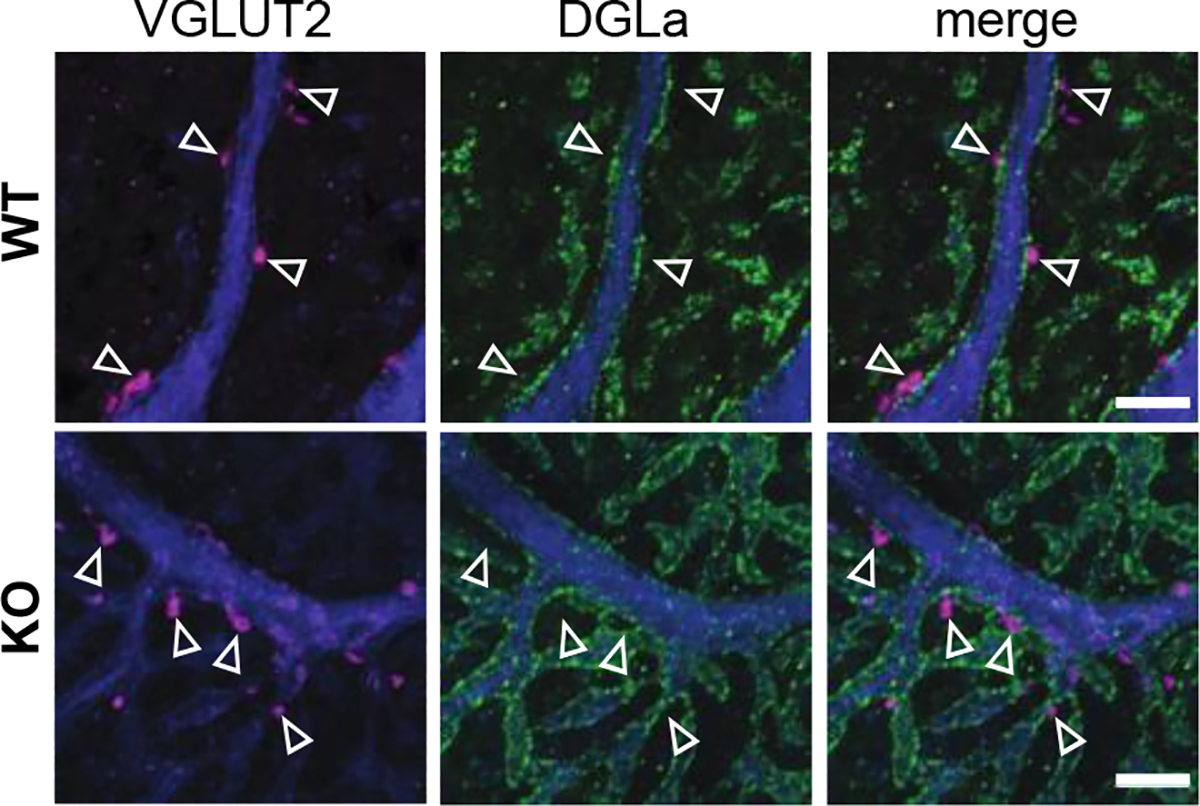
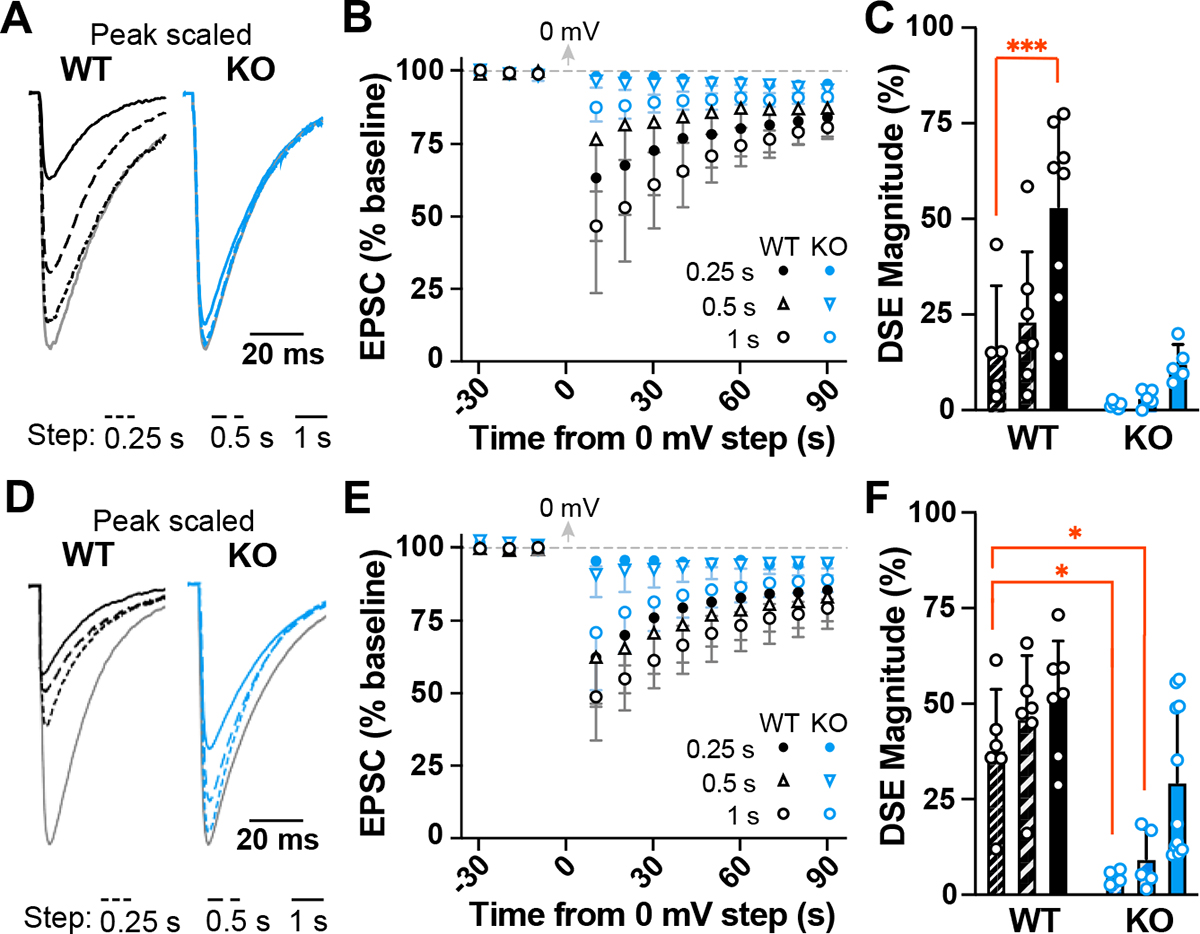
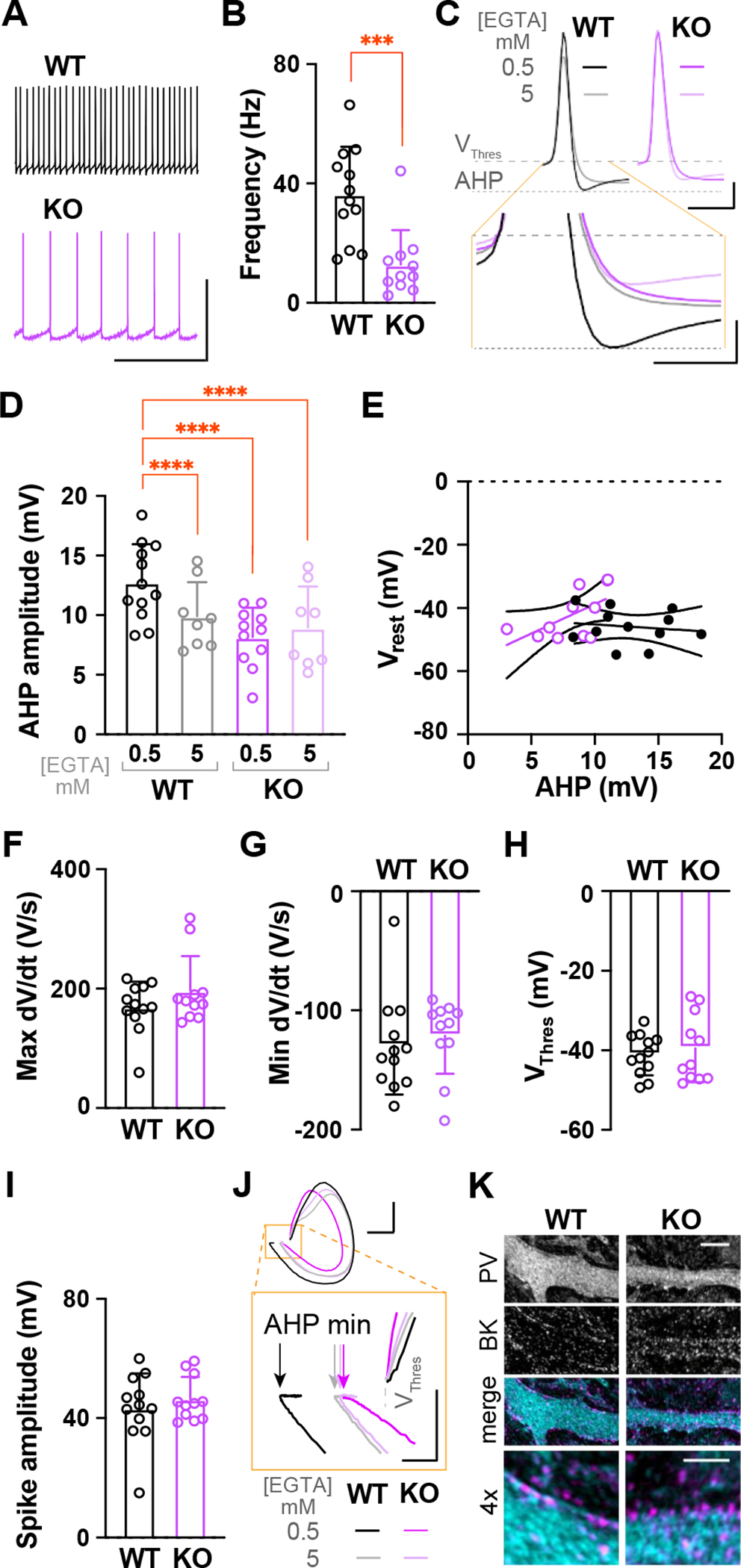
α2δ proteins (CACNA2D1-4) are required for normal neurological function and contribute to membrane trafficking of voltage-gated calcium channels, through which calcium entry initiates numerous physiological processes. However, it remains unclear how α2δ proteins influence calcium-mediated signalling to control neuronal output. Using whole-cell recordings of mouse Purkinje cells, we show that α2δ-2 is required for functional coupling of postsynaptic voltage-dependent calcium entry with calcium-dependent effector mechanisms controlling two different outputs, depolarization-induced suppression of excitation and spike afterhyperpolarization. Our findings indicate an important role for α2δ-2 proteins in regulating functional postsynaptic calcium channel coupling in neurons, providing new context for understanding the effects of α2δ mutations on neuronal circuit function and presenting additional potential avenues to manipulate α2δ-mediated signalling for therapeutic gain. KEY POINTS: Calcium influx, via voltage-dependent calcium channels, drives numerous neuronal signalling processes with precision achieved in part by tight coupling between calcium entry and calcium-dependent effectors. α2δ proteins are important for neurological function and contribute to calcium channel membrane trafficking, although how α2δ proteins influence postsynaptic calcium-dependent signalling is largely unexplored. Here it is shown that loss of α2δ-2 proteins disrupts functional calcium coupling to two different postsynaptic calcium-dependent signals in mouse Purkinje cell neurons, retrograde endocannabinoid signalling and the action potential afterhyperpolarization. The findings provide new insights into the control of calcium coupling as well as new roles for α2δ-2 proteins in neurons.
Keywords: calcium channels; depolarization-induced suppression of excitation; endocannabinoid signalling; nanodomains; purkinje cells; α2δ-2 proteins.
© 2021 The Authors. The Journal of Physiology © 2021 The Physiological Society.
Conflict of interest statement
Figures
Similar articles
-
α2δ-2 Protein Controls Structure and Function at the Cerebellar Climbing Fiber Synapse.J Neurosci. 2020 Mar 18;40(12):2403-2415. doi: 10.1523/JNEUROSCI.1514-19.2020. Epub 2020 Feb 21. J Neurosci. 2020. PMID: 32086258 Free PMC article.
-
α-Neurexins Together with α2δ-1 Auxiliary Subunits Regulate Ca2+ Influx through Cav2.1 Channels.J Neurosci. 2018 Sep 19;38(38):8277-8294. doi: 10.1523/JNEUROSCI.0511-18.2018. Epub 2018 Aug 13. J Neurosci. 2018. PMID: 30104341 Free PMC article.
-
Purine receptor-mediated endocannabinoid production and retrograde synaptic signalling in the cerebellar cortex.Br J Pharmacol. 2011 Feb;162(4):974-88. doi: 10.1111/j.1476-5381.2010.01106.x. Br J Pharmacol. 2011. PMID: 21054344 Free PMC article.
-
Neuronal α2δ proteins and brain disorders.Pflugers Arch. 2020 Jul;472(7):845-863. doi: 10.1007/s00424-020-02420-2. Epub 2020 Jun 30. Pflugers Arch. 2020. PMID: 32607809 Free PMC article. Review.
-
Voltage-dependent calcium channels.Gen Physiol Biophys. 2005 Jun;24 Suppl 1:1-78. Gen Physiol Biophys. 2005. PMID: 16096350 Review.
Cited by
-
Aberrant DJ-1 expression underlies L-type calcium channel hypoactivity in dendrites in tuberous sclerosis complex and Alzheimer's disease.Proc Natl Acad Sci U S A. 2023 Nov 7;120(45):e2301534120. doi: 10.1073/pnas.2301534120. Epub 2023 Oct 30. Proc Natl Acad Sci U S A. 2023. PMID: 37903257 Free PMC article.
-
Altered hippocampal activation in seizure-prone CACNA2D2 knockout mice.bioRxiv [Preprint]. 2023 Nov 10:2023.11.08.565511. doi: 10.1101/2023.11.08.565511. bioRxiv. 2023. Update in: eNeuro. 2024 May 15;11(5):ENEURO.0486-23.2024. doi: 10.1523/ENEURO.0486-23.2024. PMID: 37986872 Free PMC article. Updated. Preprint.
-
Altered Hippocampal Activation in Seizure-Prone CACNA2D2 Knock-out Mice.eNeuro. 2024 May 15;11(5):ENEURO.0486-23.2024. doi: 10.1523/ENEURO.0486-23.2024. Print 2024 May. eNeuro. 2024. PMID: 38749701 Free PMC article.
-
Pathophysiological Roles of Auxiliary Calcium Channel α2δ Subunits.Handb Exp Pharmacol. 2023;279:289-316. doi: 10.1007/164_2022_630. Handb Exp Pharmacol. 2023. PMID: 36598609
-
A biallelic mutation in CACNA2D2 associated with developmental and epileptic encephalopathy affects calcium channel-dependent as well as synaptic functions of α2δ-2.J Neurochem. 2025 Jan;169(1):e16197. doi: 10.1111/jnc.16197. Epub 2024 Aug 19. J Neurochem. 2025. PMID: 39161180 Free PMC article.
References
-
- Brown J & Randall A. (2005). Gabapentin Fails to Alter P/Q-type Ca2+ Channel-Mediated Synaptic Transmission in the Hippocampus In Vitro. Synapse 55, 262–269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources