

Oxidative Metabolism as a Modulator of Kratom's Biological Actions
- PMID: 34783240
- PMCID: PMC8673317
- DOI: 10.1021/acs.jmedchem.1c01111
Oxidative Metabolism as a Modulator of Kratom's Biological Actions
Abstract
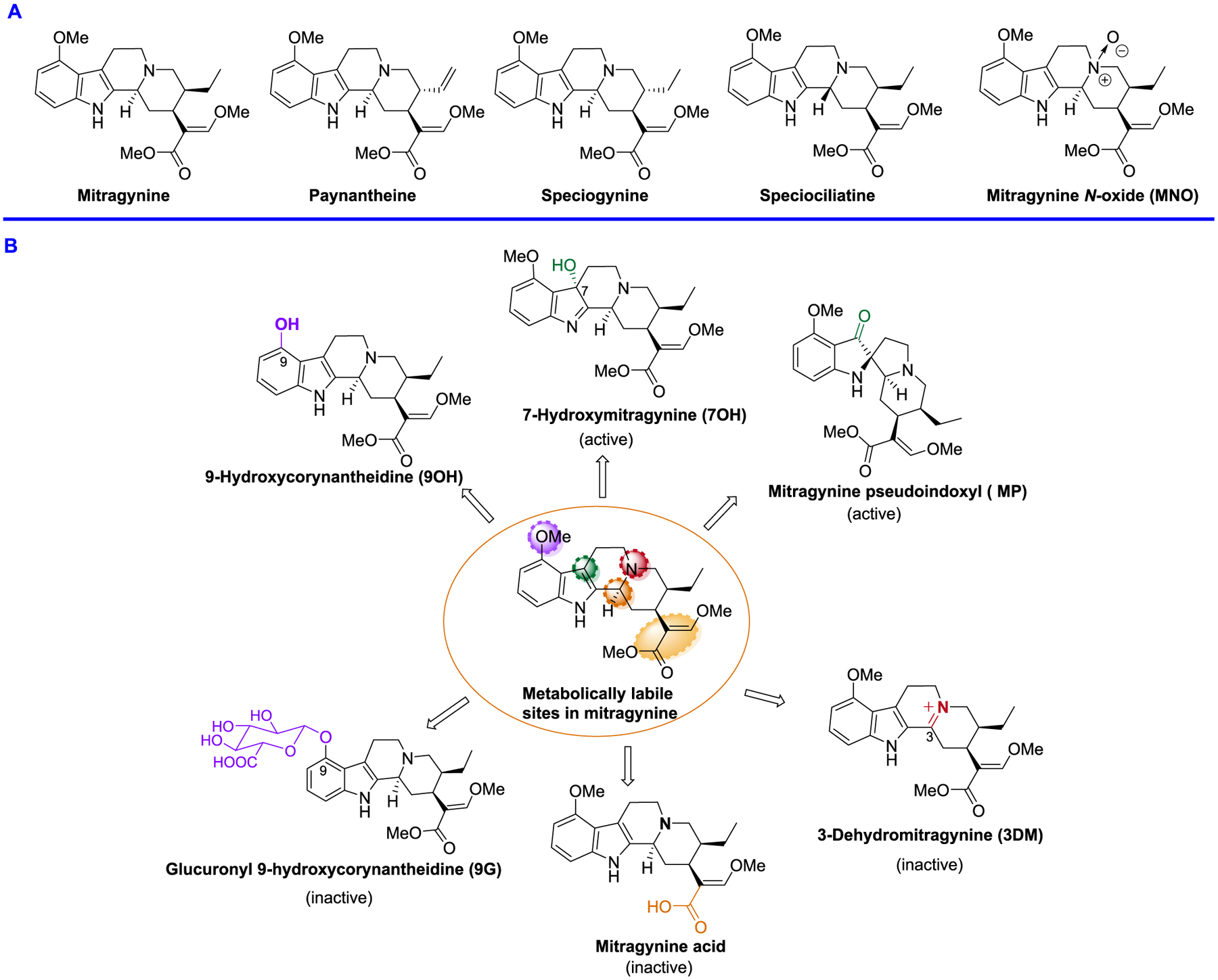
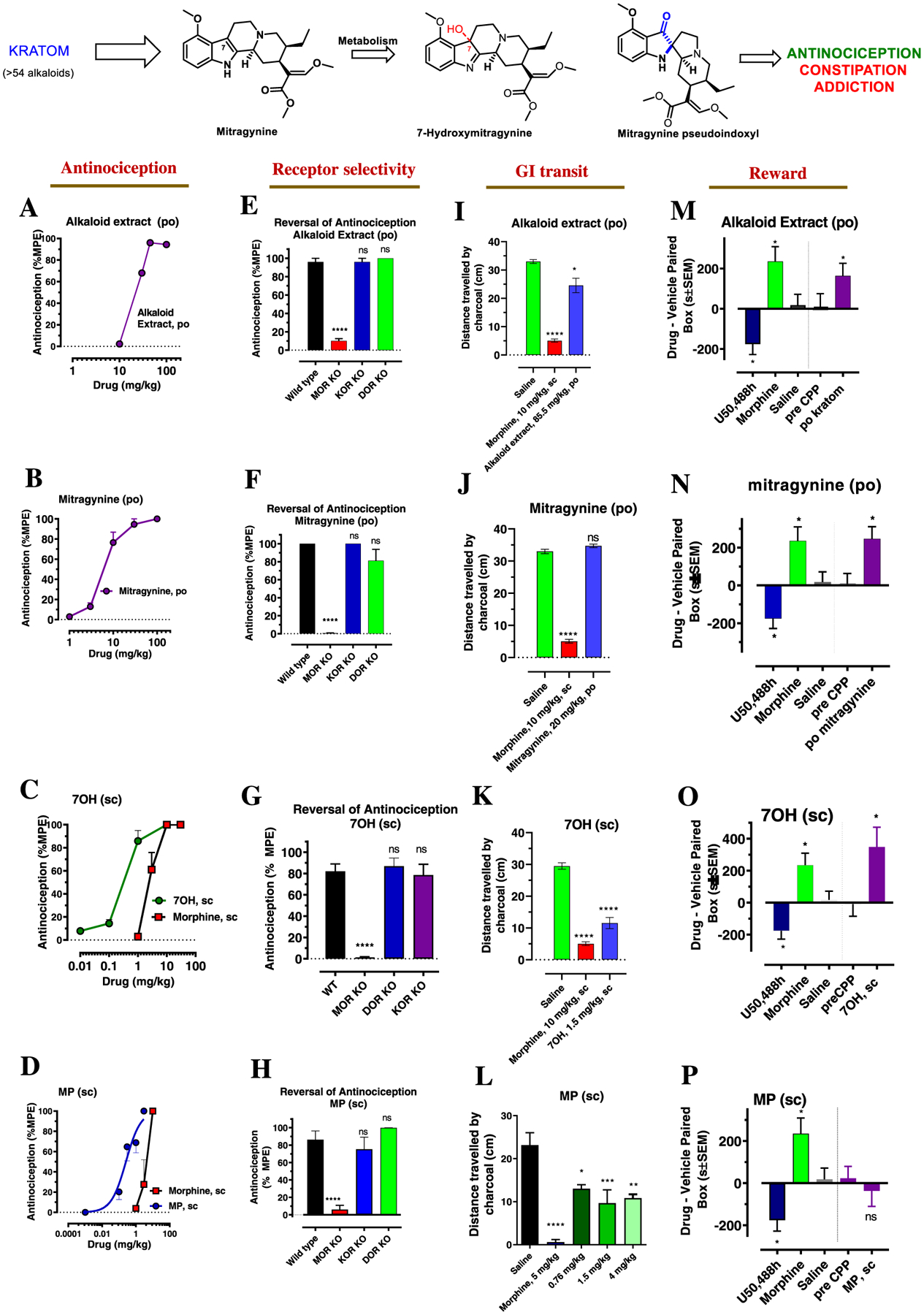
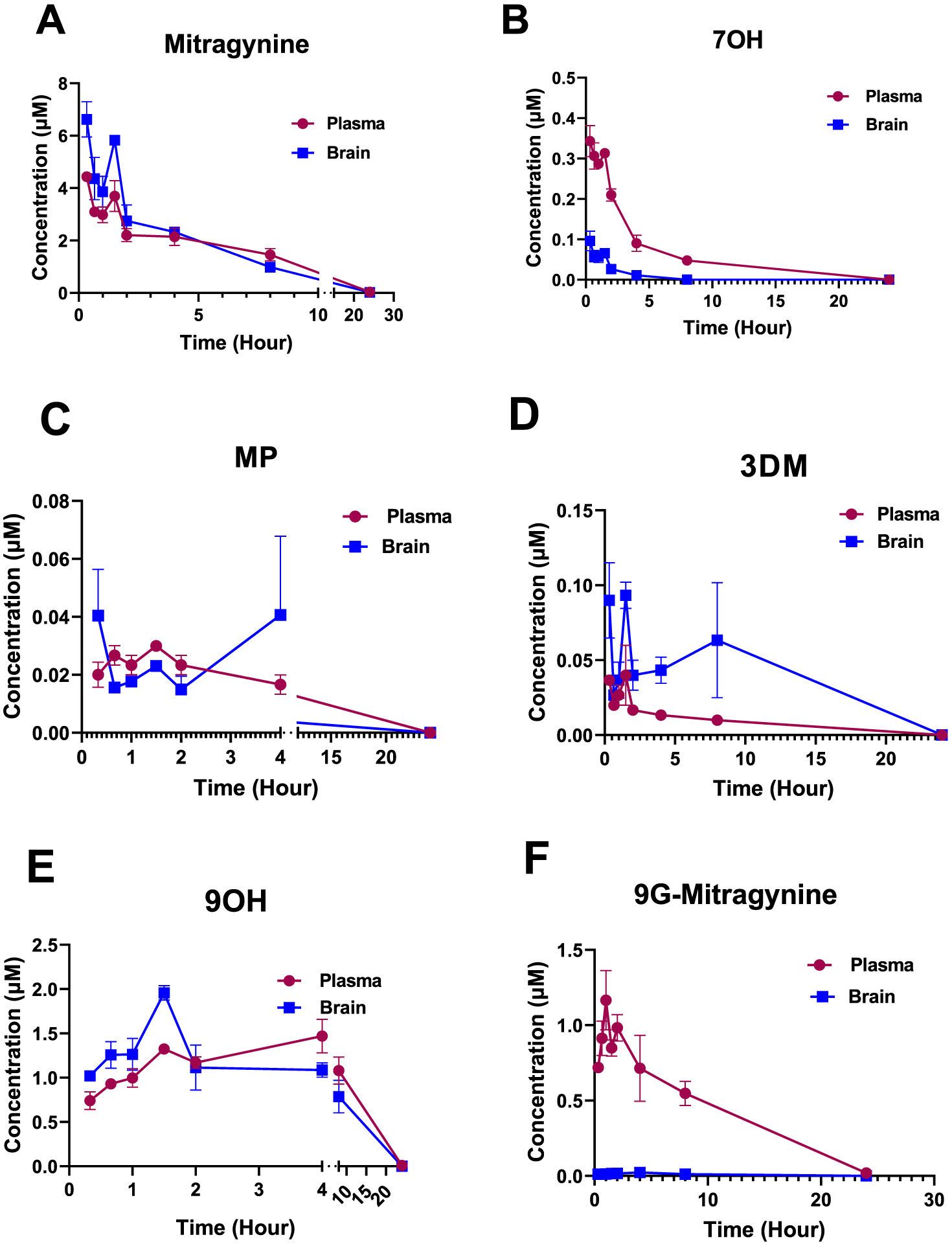
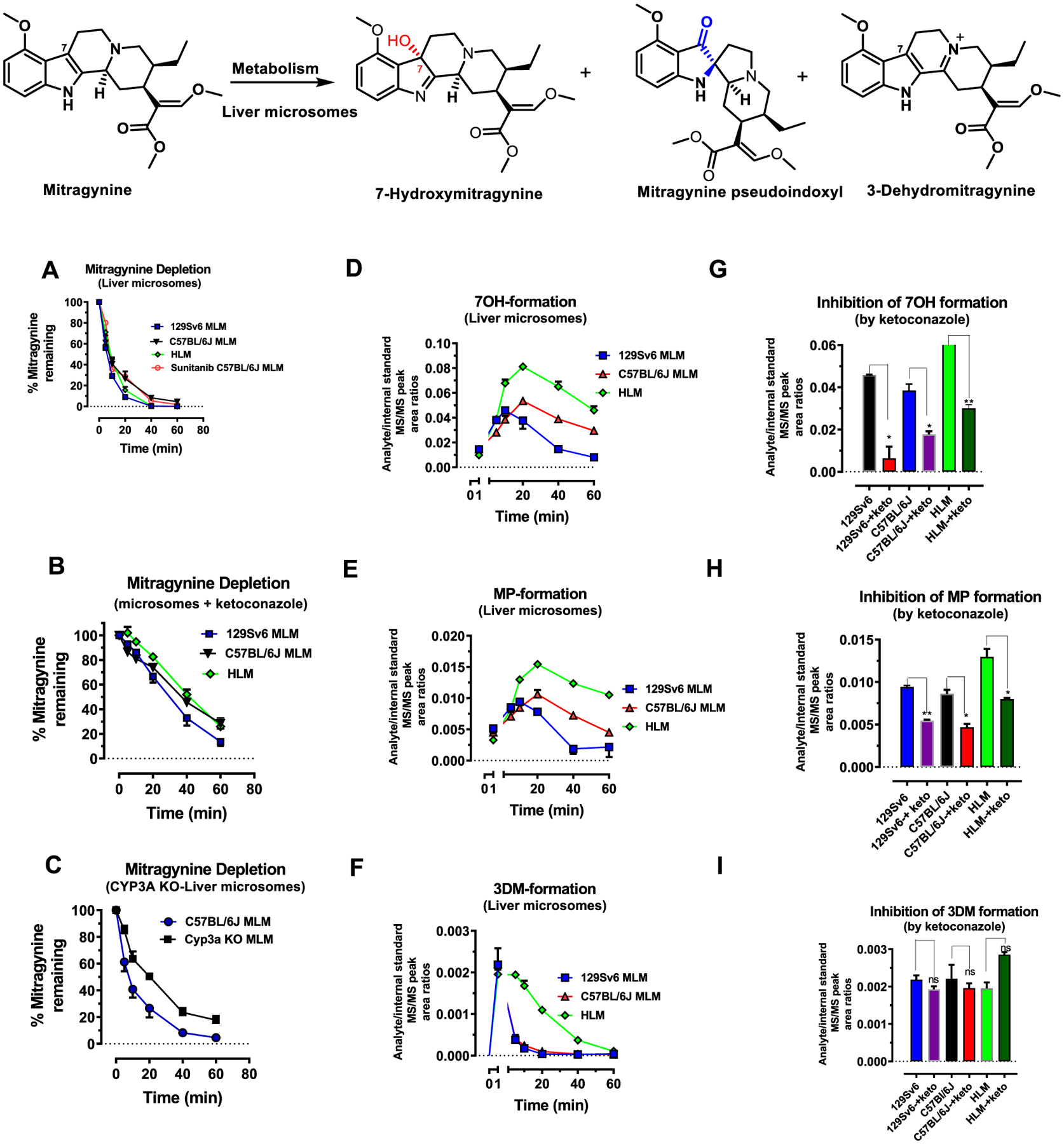
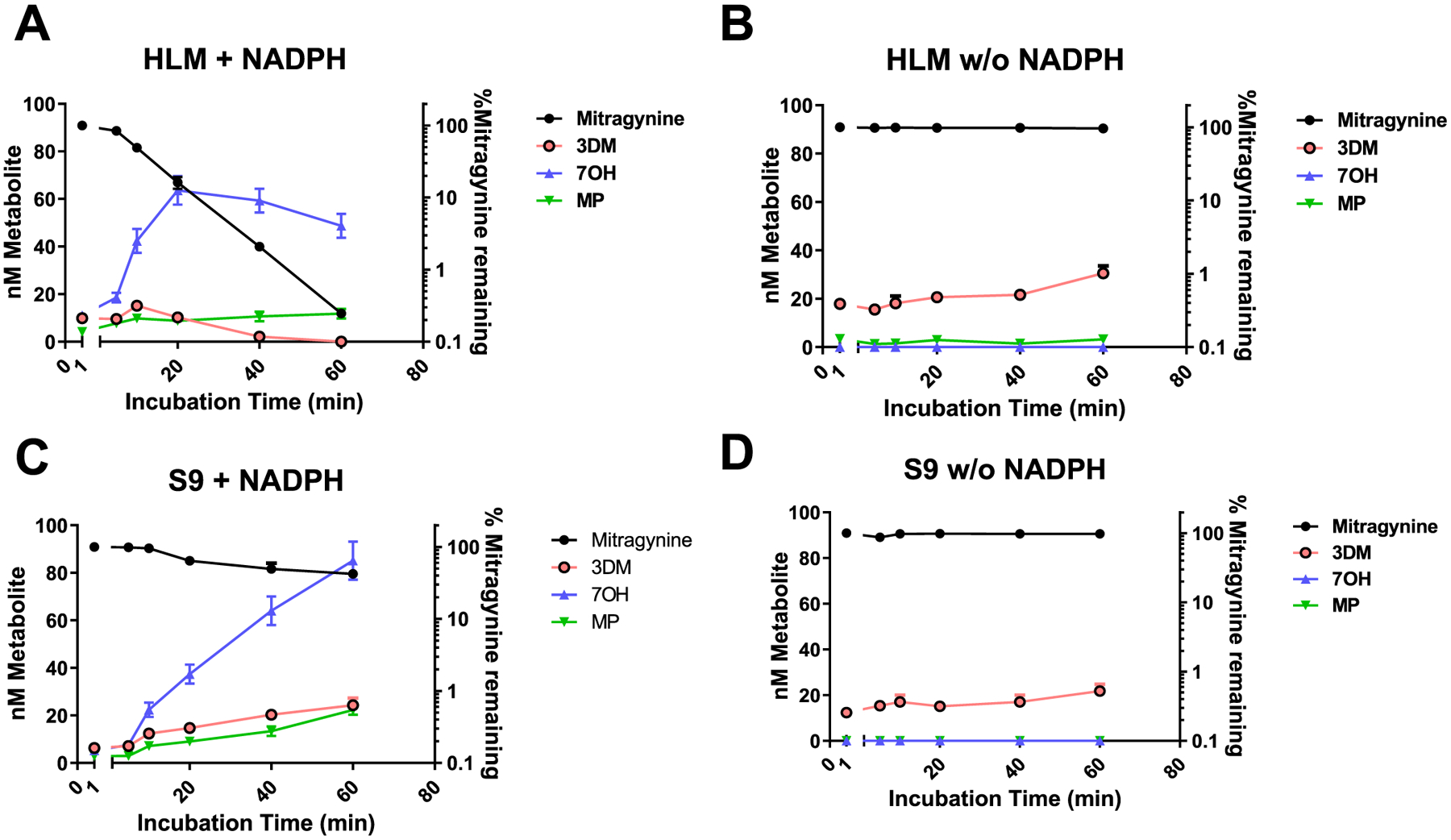
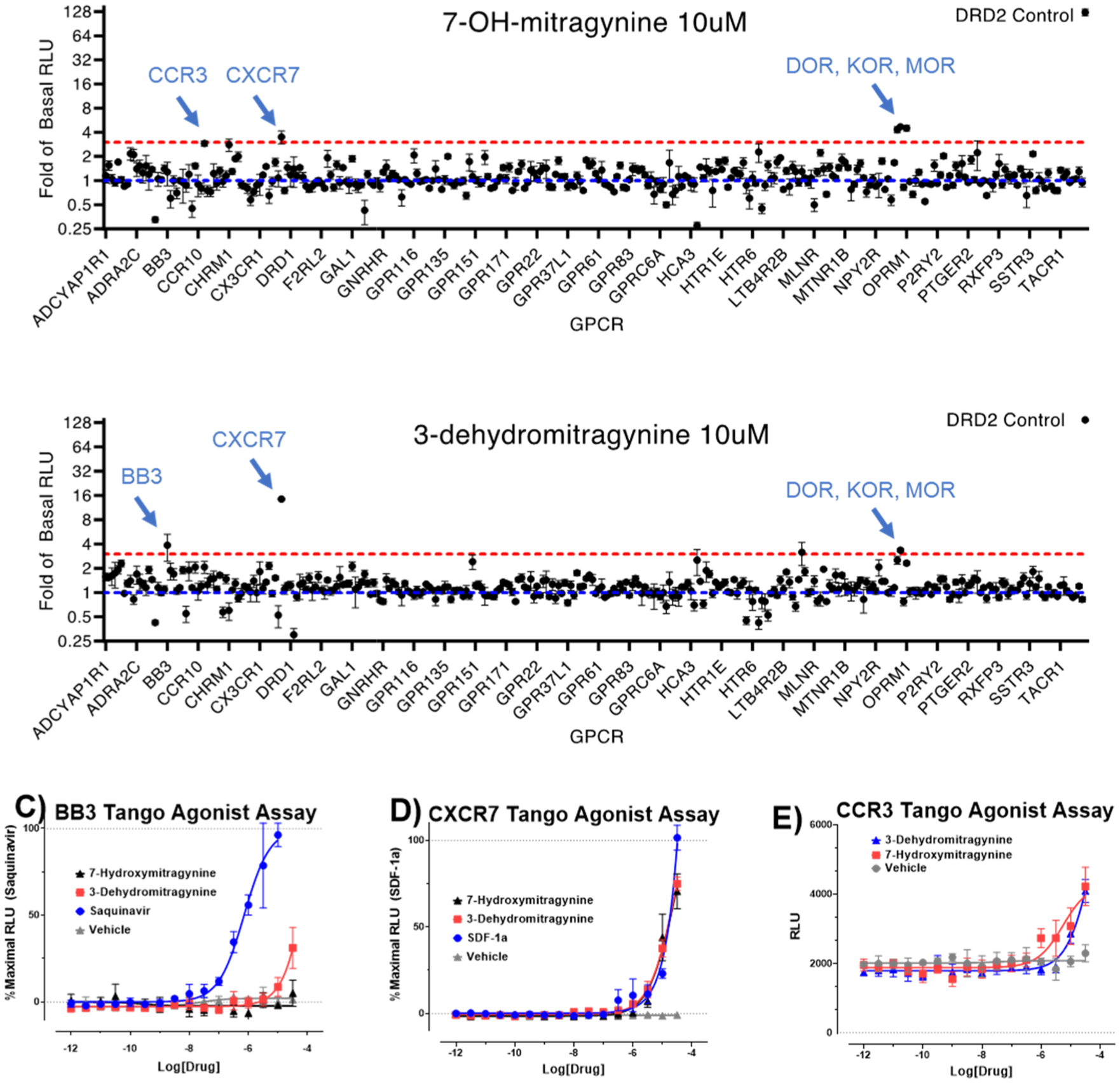
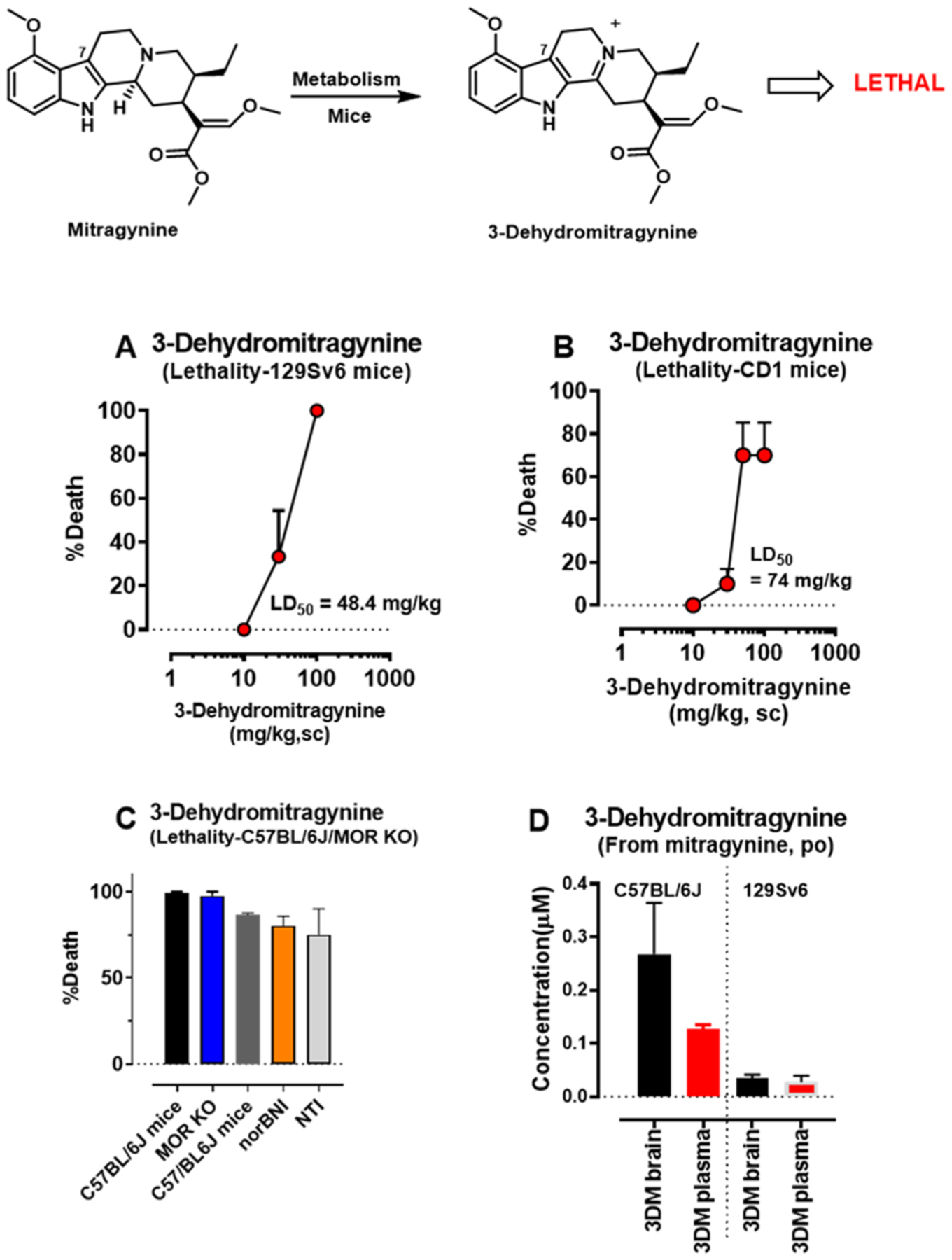
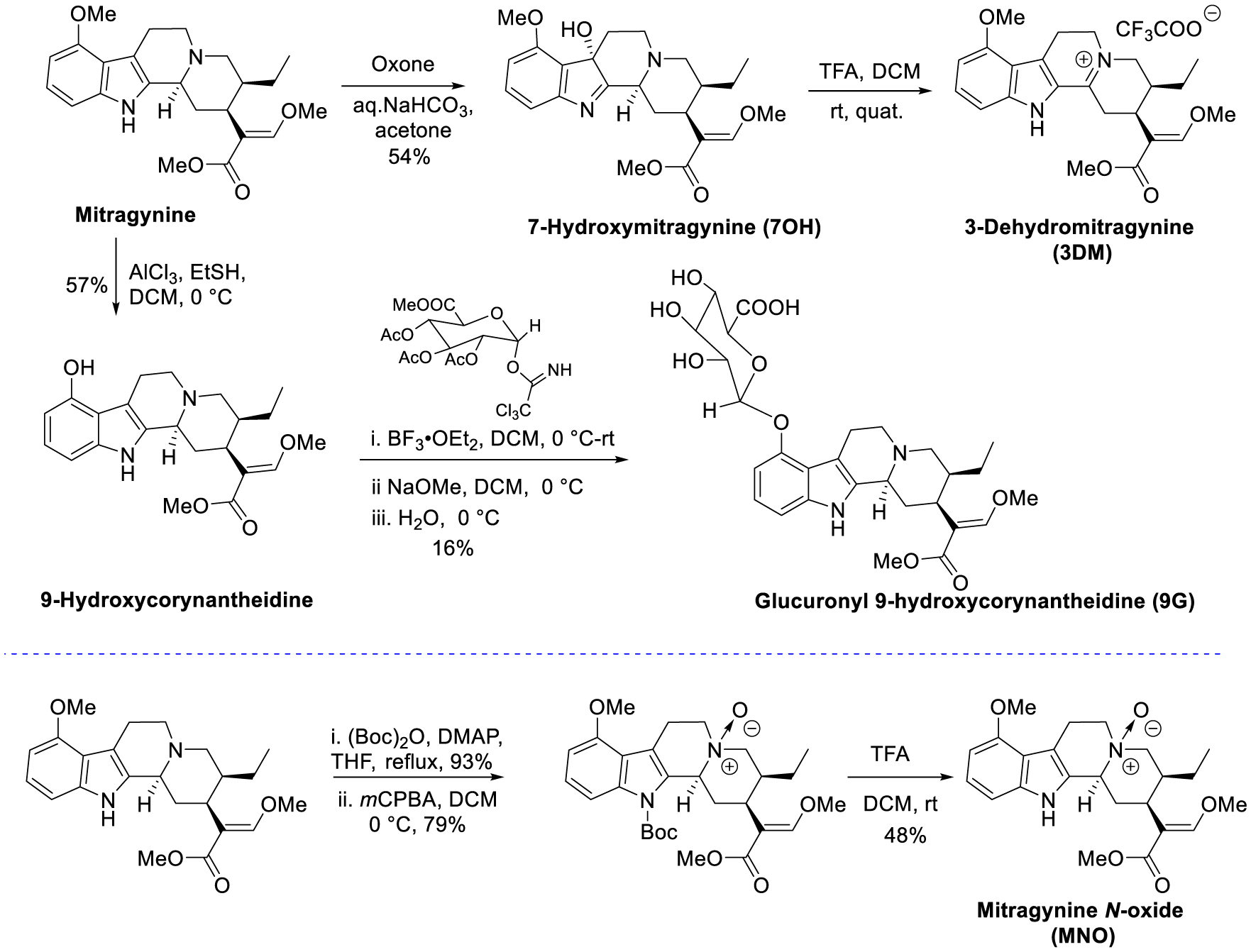
The leaves of Mitragyna speciosa (kratom), a plant native to Southeast Asia, are increasingly used as a pain reliever and for attenuation of opioid withdrawal symptoms. Using the tools of natural products chemistry, chemical synthesis, and pharmacology, we provide a detailed in vitro and in vivo pharmacological characterization of the alkaloids in kratom. We report that metabolism of kratom's major alkaloid, mitragynine, in mice leads to formation of (a) a potent mu opioid receptor agonist antinociceptive agent, 7-hydroxymitragynine, through a CYP3A-mediated pathway, which exhibits reinforcing properties, inhibition of gastrointestinal (GI) transit and reduced hyperlocomotion, (b) a multifunctional mu agonist/delta-kappa antagonist, mitragynine pseudoindoxyl, through a CYP3A-mediated skeletal rearrangement, displaying reduced hyperlocomotion, inhibition of GI transit and reinforcing properties, and (c) a potentially toxic metabolite, 3-dehydromitragynine, through a non-CYP oxidation pathway. Our results indicate that the oxidative metabolism of the mitragynine template beyond 7-hydroxymitragynine may have implications in its overall pharmacology in vivo.
Conflict of interest statement
The authors declare the following competing financial interest(s): SM, GWP, YXP are co-founders of Sparian biosciences. DS and ACK are co-founders of Kures, Inc. GWP, ACK, DS, AK, AV and SM have filed a provisional patents on mitragynine and related molecules. The other authors declare no other competing interests.
Figures
References
-
- Institute of Medicine (US) Committee on Advancing Pain Research, C. and E. Relieving Pain in America: A Blueprint for Transforming Prevention, Care, Education, and Research; National Academies Press, 2011. - PubMed
-
- Imam MZ; Kuo A; Ghassabian S; Smith MT Progress in Understanding Mechanisms of Opioid-Induced Gastrointestinal Adverse Effects and Respiratory Depression. Neuropharmacology 2018, 131, 238–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Chemical Information
Research Materials
