

Systematic exploration of Escherichia coli phage-host interactions with the BASEL phage collection
- PMID: 34784345
- PMCID: PMC8594841
- DOI: 10.1371/journal.pbio.3001424
Systematic exploration of Escherichia coli phage-host interactions with the BASEL phage collection
Abstract
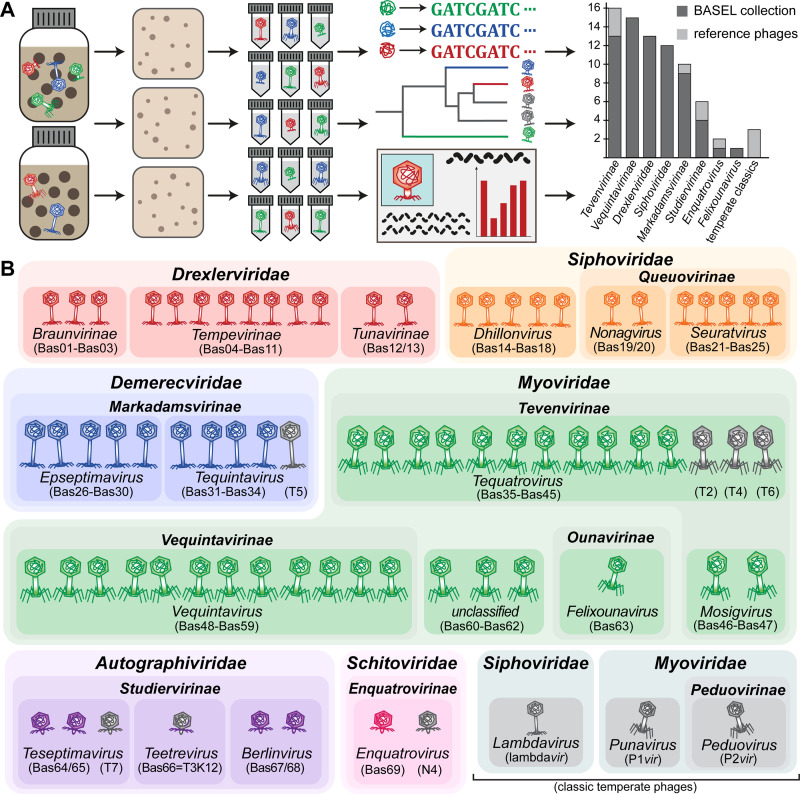
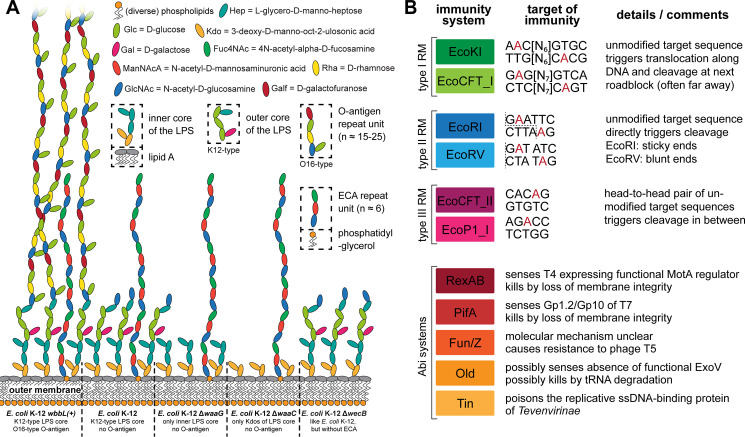
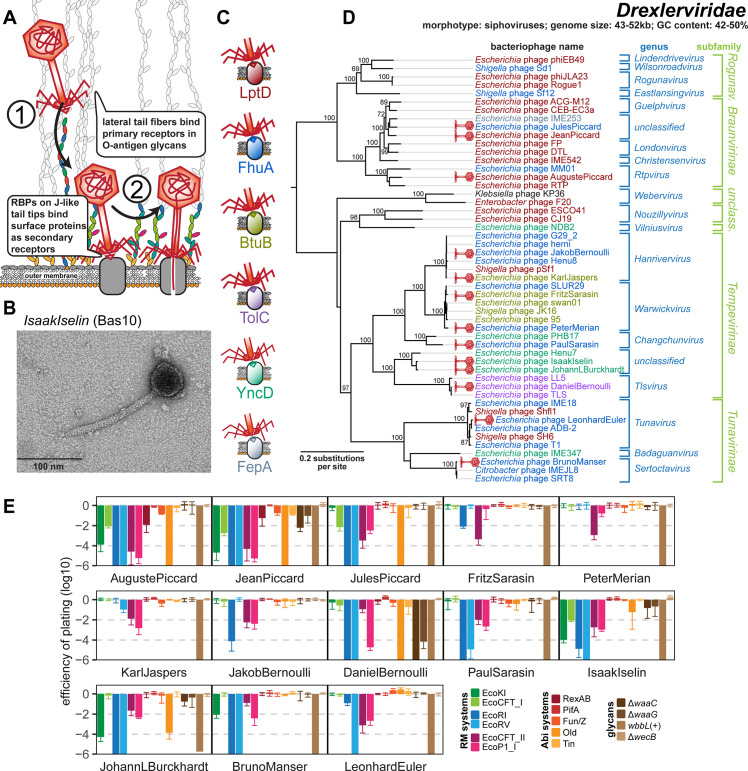
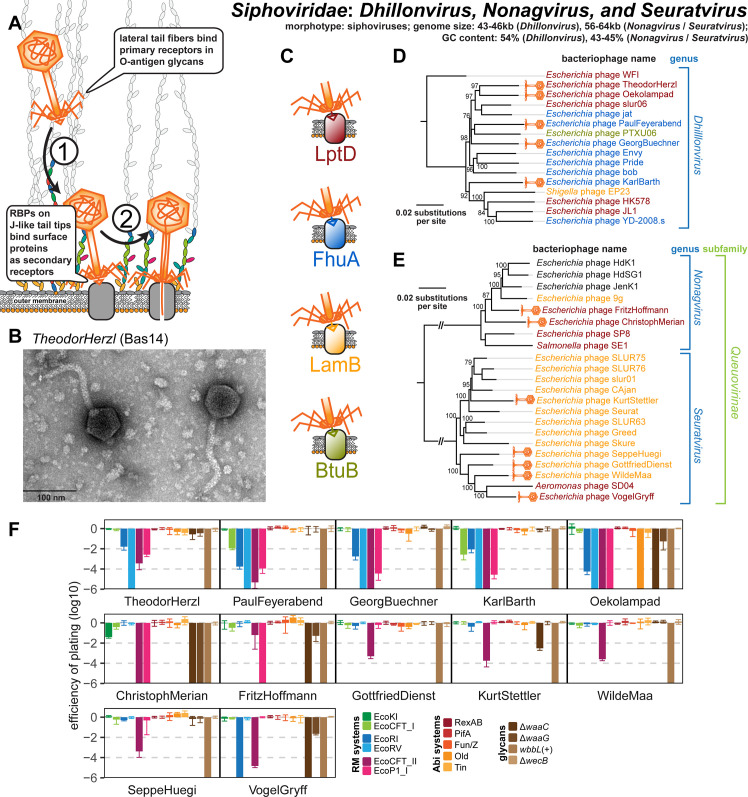
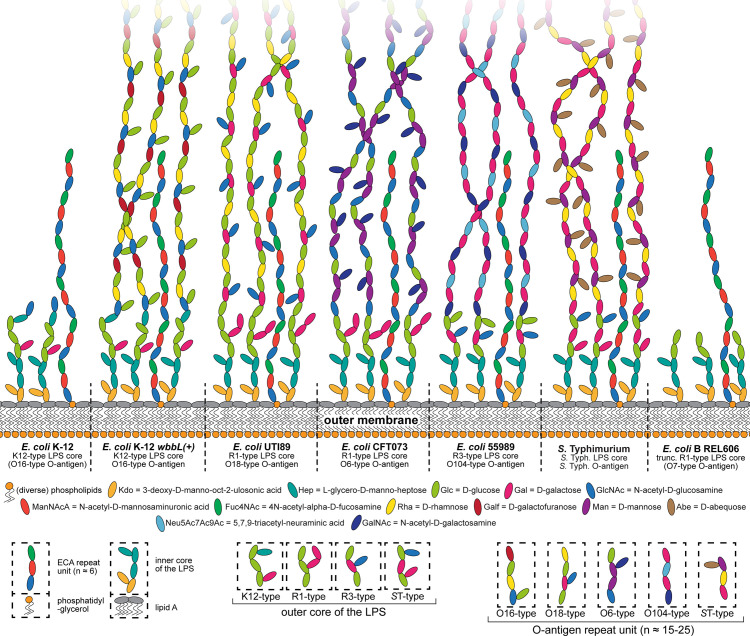
Bacteriophages, the viruses infecting bacteria, hold great potential for the treatment of multidrug-resistant bacterial infections and other applications due to their unparalleled diversity and recent breakthroughs in their genetic engineering. However, fundamental knowledge of the molecular mechanisms underlying phage-host interactions is mostly confined to a few traditional model systems and did not keep pace with the recent massive expansion of the field. The true potential of molecular biology encoded by these viruses has therefore remained largely untapped, and phages for therapy or other applications are often still selected empirically. We therefore sought to promote a systematic exploration of phage-host interactions by composing a well-assorted library of 68 newly isolated phages infecting the model organism Escherichia coli that we share with the community as the BASEL (BActeriophage SElection for your Laboratory) collection. This collection is largely representative of natural E. coli phage diversity and was intensively characterized phenotypically and genomically alongside 10 well-studied traditional model phages. We experimentally determined essential host receptors of all phages, quantified their sensitivity to 11 defense systems across different layers of bacterial immunity, and matched these results to the phages' host range across a panel of pathogenic enterobacterial strains. Clear patterns in the distribution of phage phenotypes and genomic features highlighted systematic differences in the potency of different immunity systems and suggested the molecular basis of receptor specificity in several phage groups. Our results also indicate strong trade-offs between fitness traits like broad host recognition and resistance to bacterial immunity that might drive the divergent adaptation of different phage groups to specific ecological niches. We envision that the BASEL collection will inspire future work exploring the biology of bacteriophages and their hosts by facilitating the discovery of underlying molecular mechanisms as the basis for an effective translation into biotechnology or therapeutic applications.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

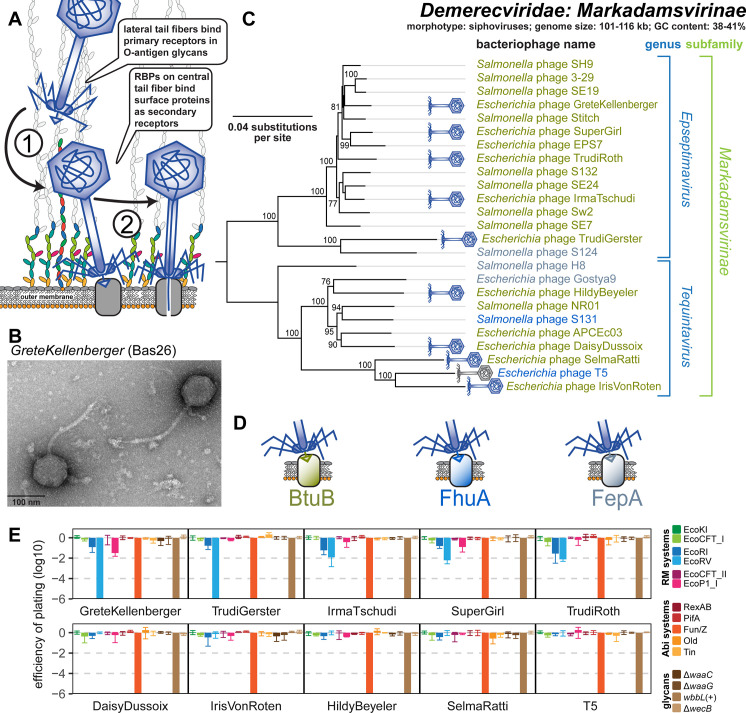

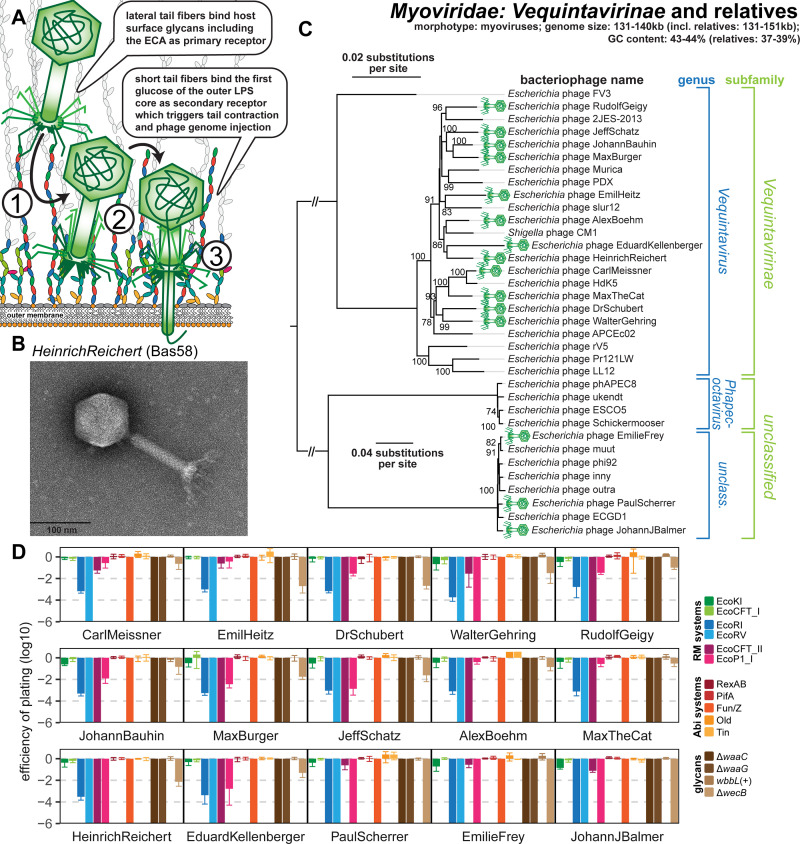
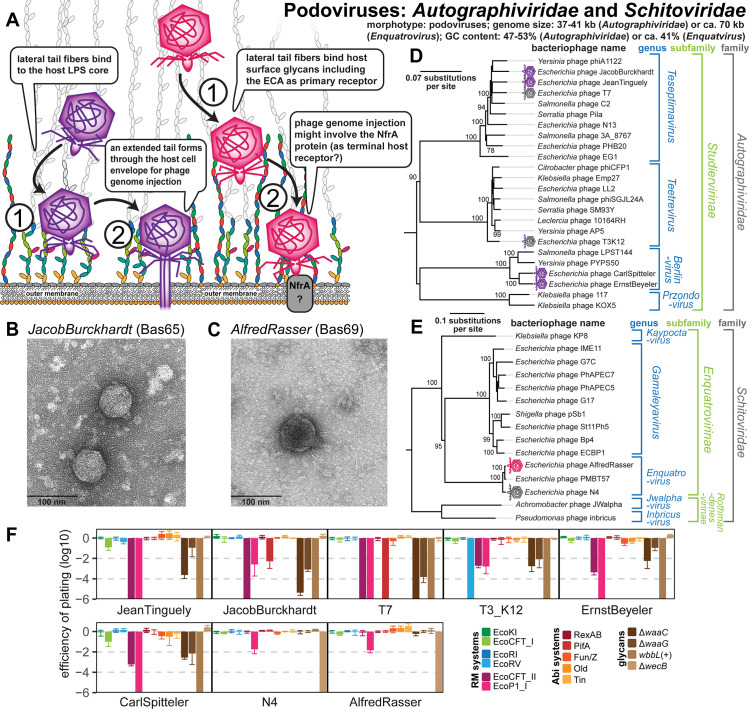
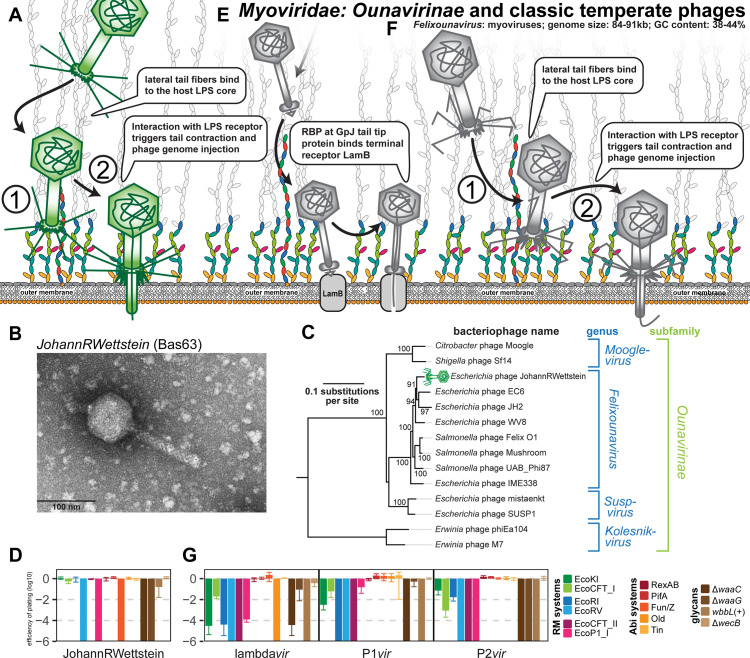

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
