

Deletion of p16 accelerates fracture healing in geriatric mice
- PMID: 34786046
- PMCID: PMC8581914
Deletion of p16 accelerates fracture healing in geriatric mice
Abstract
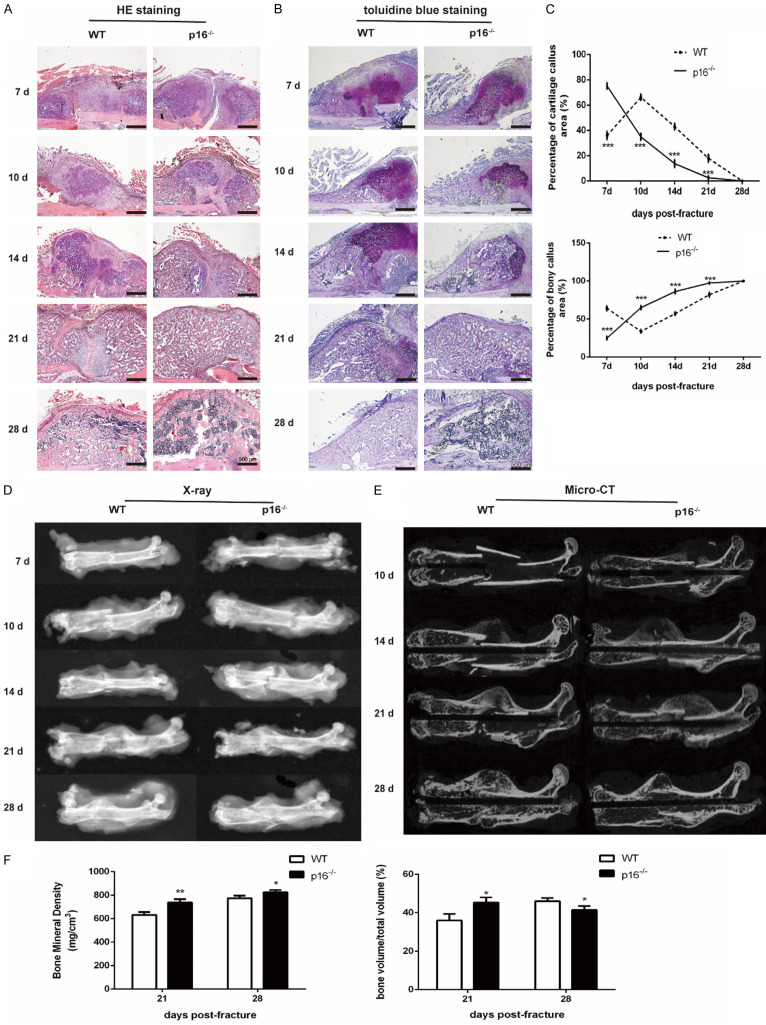
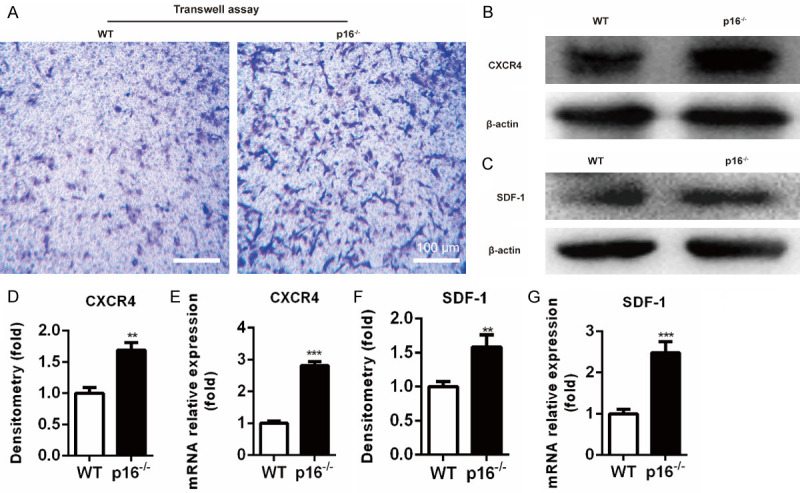
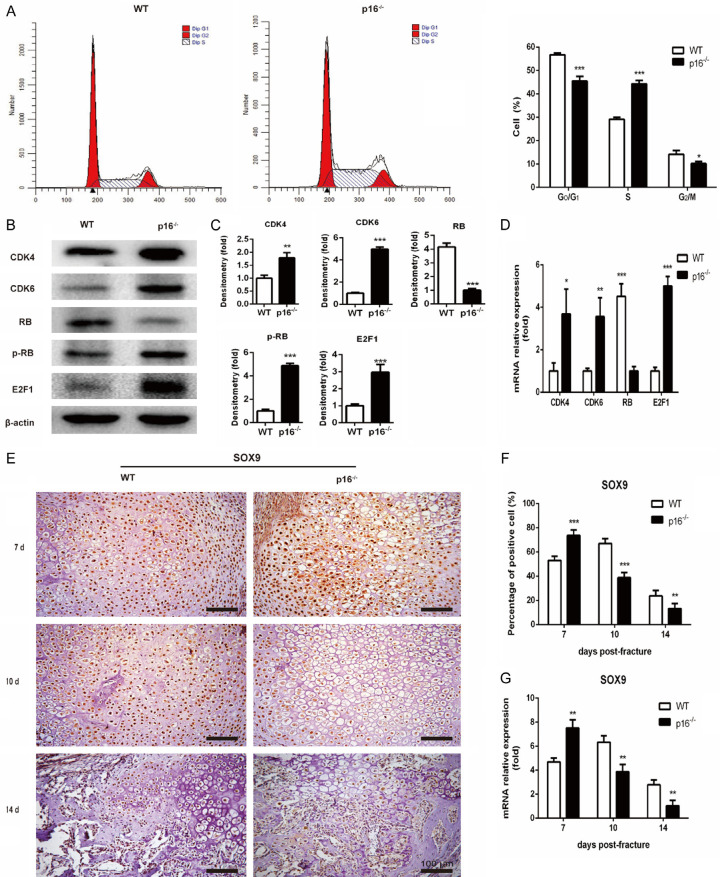
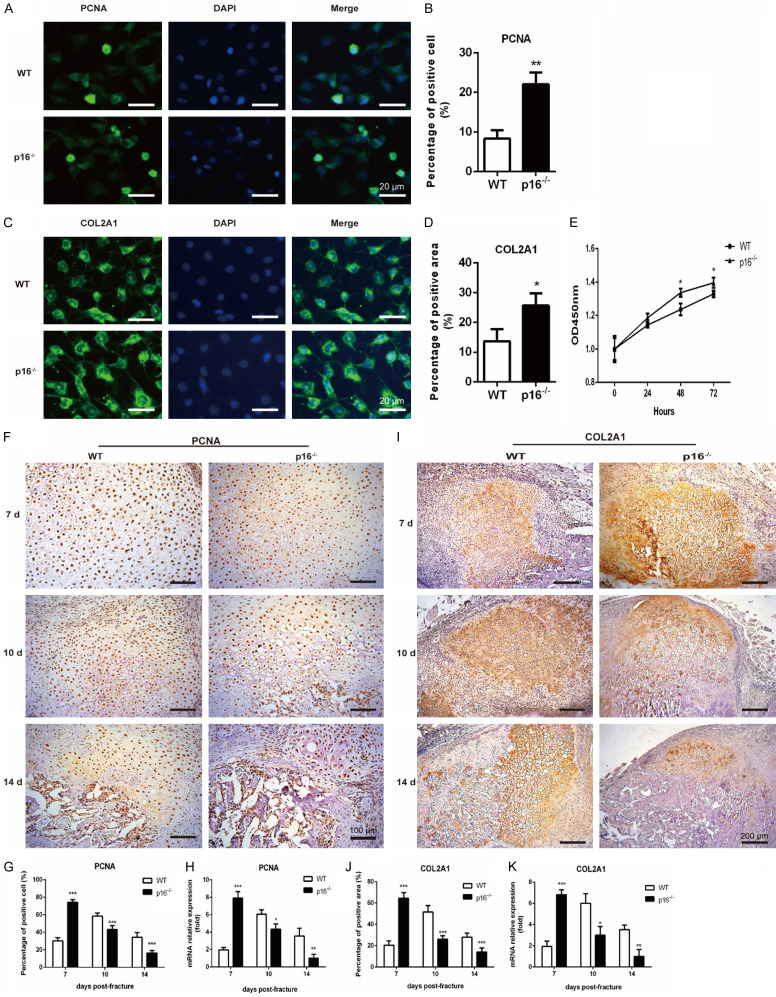
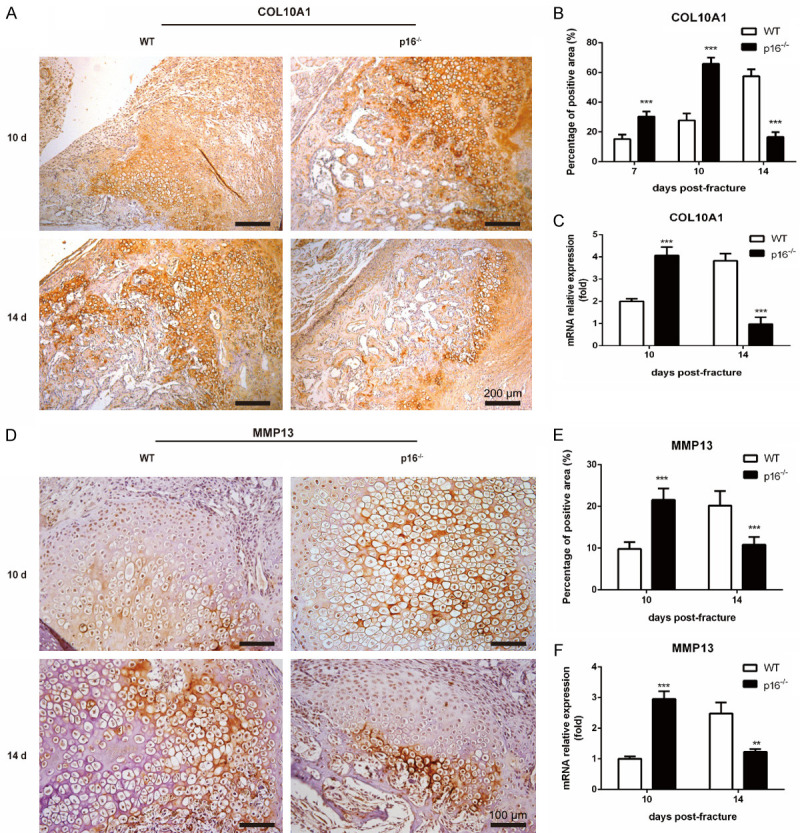
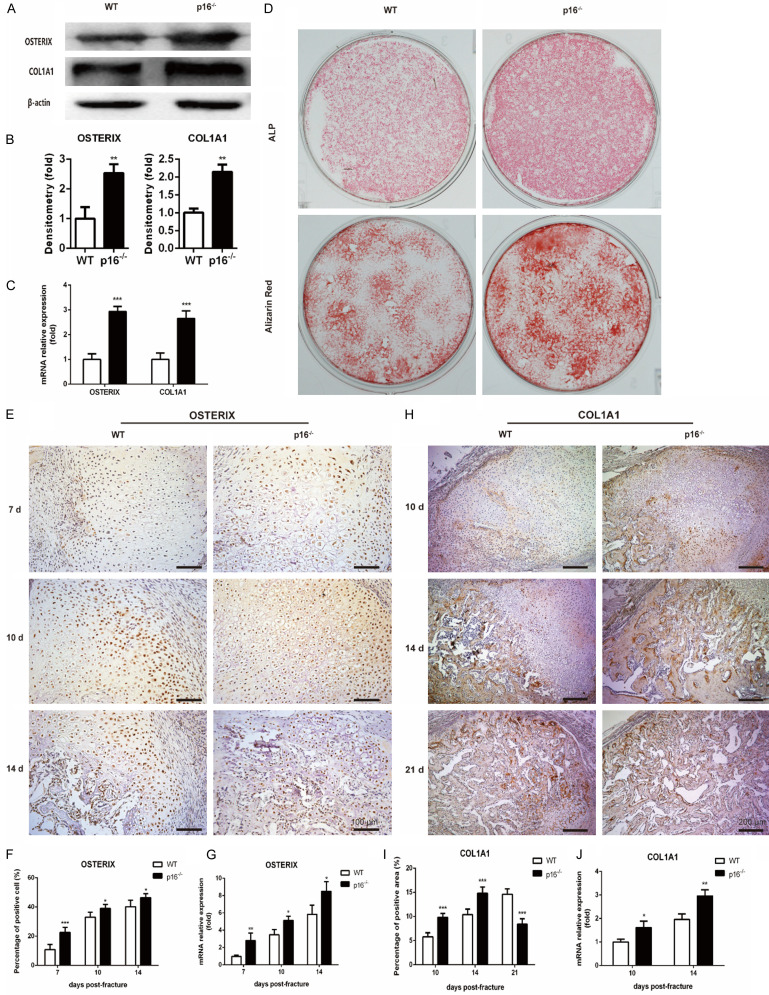
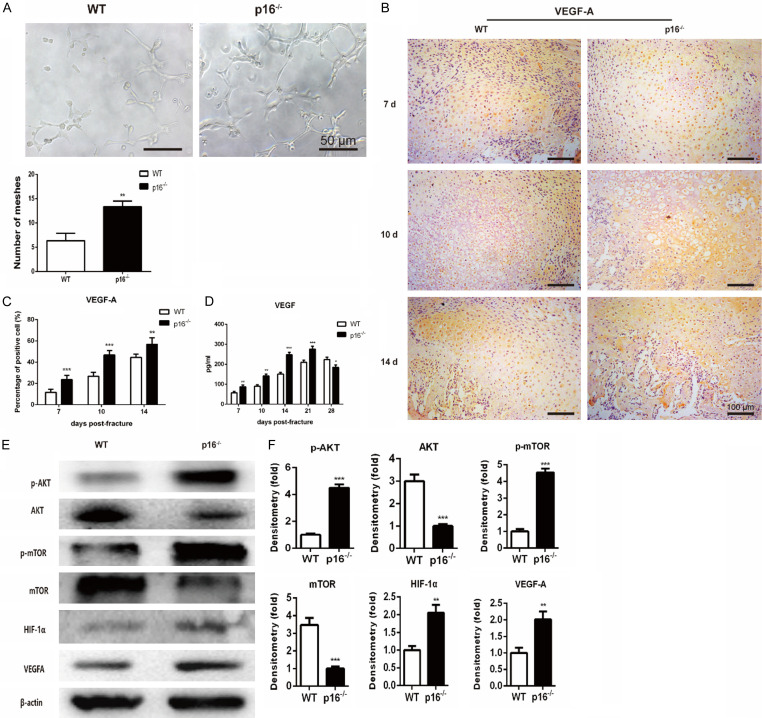
The biomarker p16 plays a role in aging and is upregulated in aged organs and cells, including bone marrow mesenchymal stem cells (BM-MSCs), which play a leading role in fracture healing. Several studies have reported delayed fracture healing in geriatric mice. However, the relationship between p16 expression and fracture healing in geriatric mice remains poorly understood. In this study, we found that fracture healing was accelerated in p16 deletion (p16-/-) mice, and the number of migrated BM-MSCs from p16-/- mice increased. The expressions of SDF-1 and CXCR4 were also upregulated in p16-/- mice. Increased cell percentage at S phase in cell cycle, enhanced expressions of CDK4/6, pRB, and E2F1, decreased expression of RB, and elevated expressions of SOX9, PCNA, and COL2A1 were detected in p16-/- mice. The expressions of COL10A1, MMP13, OSTERIX, and COL1A1 were also high in p16-/- mice. Moreover, the expressions of p-AKT, p-mTOR, HIF-1α, and VEGF-A in BM-MSCs and expression of VEGF-A in callus were upregulated in p16-/- mice. The expression of VEGF in the serum of p16-/- mice was also higher than that of wild type mice. Thus, deletion of p16 enhances migration, division, and differentiation of BM-MSCs, promotes proliferation and maturation of chondrocytes, activates osteoblastogenesis, and facilitates vascularization to accelerate fracture healing, providing a novel strategy to treat fracture in the elderly.
Keywords: endochondral ossification; fracture healing; geriatric mice; p16; vascularization.
AJTR Copyright © 2021.
Conflict of interest statement
None.
Figures
Similar articles
-
Aging periosteal progenitor cells have reduced regenerative responsiveness to bone injury and to the anabolic actions of PTH 1-34 treatment.Bone. 2014 May;62:79-89. doi: 10.1016/j.bone.2014.02.002. Epub 2014 Feb 12. Bone. 2014. PMID: 24530870 Free PMC article.
-
Thrombospondin-2 influences the proportion of cartilage and bone during fracture healing.J Bone Miner Res. 2009 Jun;24(6):1043-54. doi: 10.1359/jbmr.090101. J Bone Miner Res. 2009. PMID: 19123916 Free PMC article.
-
Co-culture of the bone and bone marrow: a novel way to obtain mesenchymal stem cells with enhanced osteogenic ability for fracture healing in SD rats.J Orthop Surg Res. 2019 Sep 3;14(1):293. doi: 10.1186/s13018-019-1346-z. J Orthop Surg Res. 2019. PMID: 31481070 Free PMC article.
-
Tranilast stimulates endochondral ossification by upregulating SOX9 and RUNX2 promoters.Biochem Biophys Res Commun. 2016 Feb 5;470(2):356-361. doi: 10.1016/j.bbrc.2016.01.044. Epub 2016 Jan 9. Biochem Biophys Res Commun. 2016. PMID: 26777999
-
Fracture healing in osteoporotic bone.Injury. 2016 Jun;47 Suppl 2:S21-6. doi: 10.1016/S0020-1383(16)47004-X. Injury. 2016. PMID: 27338222 Review.
Cited by
-
p16 Expression in Laryngeal Squamous Cell Carcinoma: A Surrogate or Independent Prognostic Marker?Pathogens. 2024 Jan 24;13(2):100. doi: 10.3390/pathogens13020100. Pathogens. 2024. PMID: 38392838 Free PMC article. Review.
-
Downregulation of miR-221-3p by LncRNA TUG1 Promoting the Healing of Closed Tibial Fractures in Mice.Biomed Res Int. 2022 Aug 26;2022:1624446. doi: 10.1155/2022/1624446. eCollection 2022. Biomed Res Int. 2022. Retraction in: Biomed Res Int. 2024 Feb 12;2024:9823060. doi: 10.1155/2024/9823060. PMID: 36060124 Free PMC article. Retracted.
-
P16INK4A-More Than a Senescence Marker.Life (Basel). 2022 Aug 28;12(9):1332. doi: 10.3390/life12091332. Life (Basel). 2022. PMID: 36143369 Free PMC article. Review.
-
Molecular signatures distinguish senescent cells from inflammatory cells in aged mouse callus stromal cells.Front Endocrinol (Lausanne). 2023 Feb 16;14:1090049. doi: 10.3389/fendo.2023.1090049. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36875448 Free PMC article.
-
Age-related secretion of grancalcin by macrophages induces skeletal stem/progenitor cell senescence during fracture healing.Bone Res. 2024 Jan 25;12(1):6. doi: 10.1038/s41413-023-00309-1. Bone Res. 2024. PMID: 38267422 Free PMC article.
References
-
- Gerstenfeld LC, Cullinane DM, Barnes GL, Graves DT, Einhorn TA. Fracture healing as a post-natal developmental process: molecular, spatial, and temporal aspects of its regulation. J Cell Biochem. 2003;88:873–884. - PubMed
-
- Borrelli J Jr, Pape C, Hak D, Hsu J, Lin S, Giannoudis P, Lane J. Physiological challenges of bone repair. J Orthop Trauma. 2012;26:708–711. - PubMed
-
- Meyer RA Jr, Tsahakis PJ, Martin DF, Banks DM, Harrow ME, Kiebzak GM. Age and ovariectomy impair both the normalization of mechanical properties and the accretion of mineral by the fracture callus in rats. J Orthop Res. 2001;19:428–435. - PubMed
-
- Meyer RA Jr, Meyer MH, Tenholder M, Wondracek S, Wasserman R, Garges P. Gene expression in older rats with delayed union of femoral fractures. J Bone Joint Surg Am. 2003;85:1243–1254. - PubMed
-
- Bak B, Andreassen TT. The effect of aging on fracture healing in the rat. Calcif Tissue Int. 1989;45:292–297. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous