

A molecular toolbox for ADP-ribosyl binding proteins
- PMID: 34786571
- PMCID: PMC8580838
- DOI: 10.1016/j.crmeth.2021.100121
A molecular toolbox for ADP-ribosyl binding proteins
Abstract
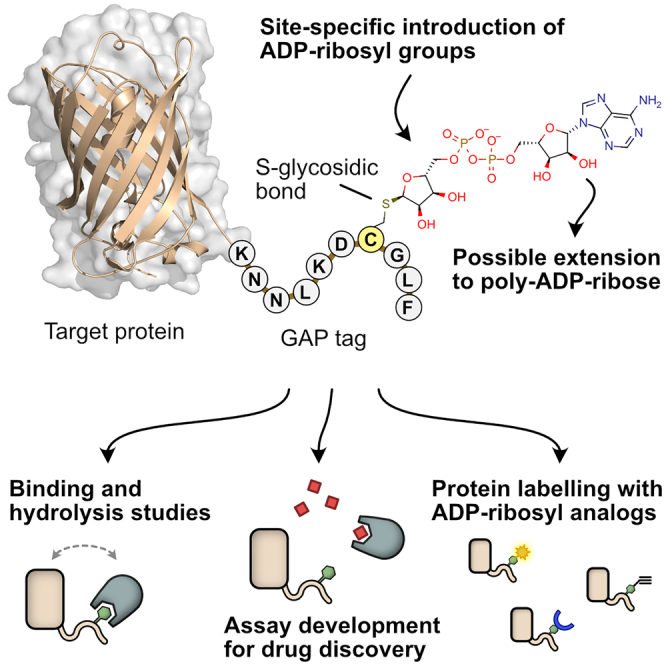
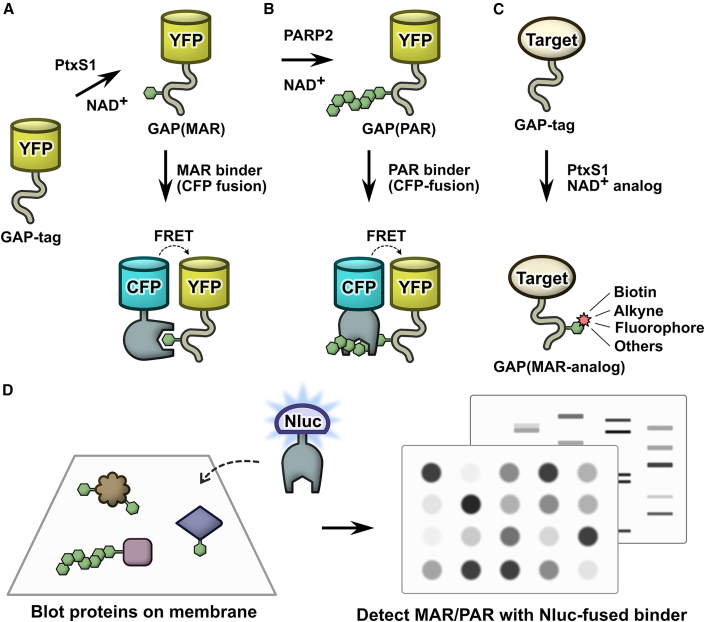
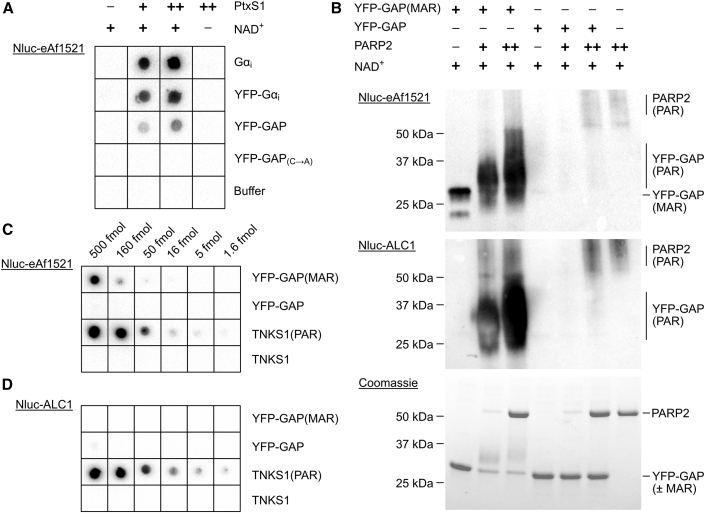
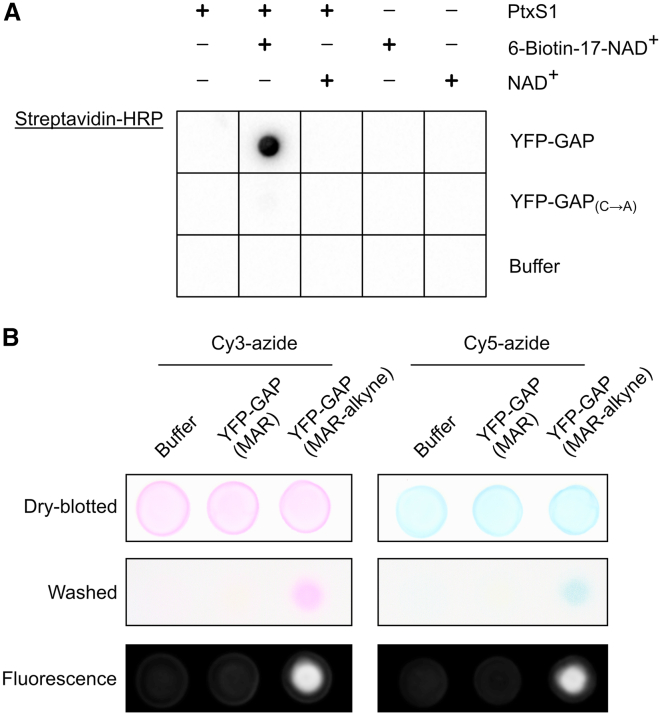
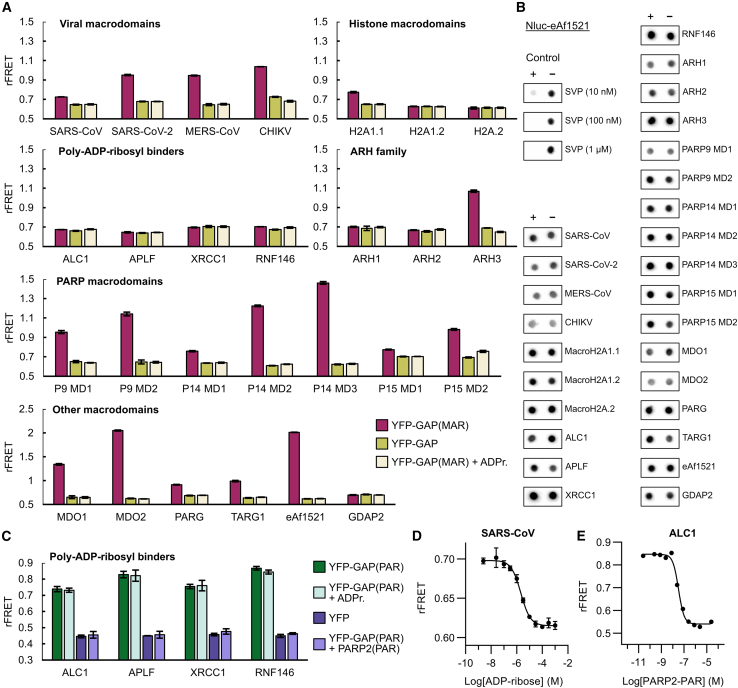
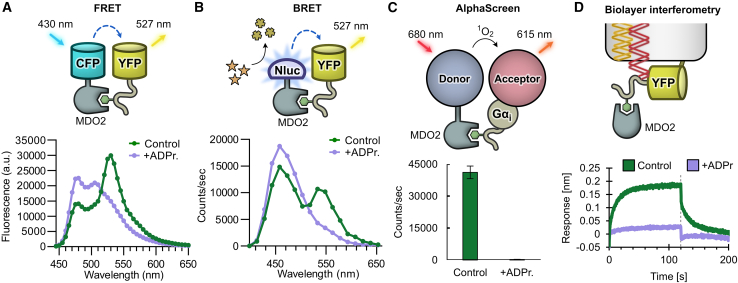
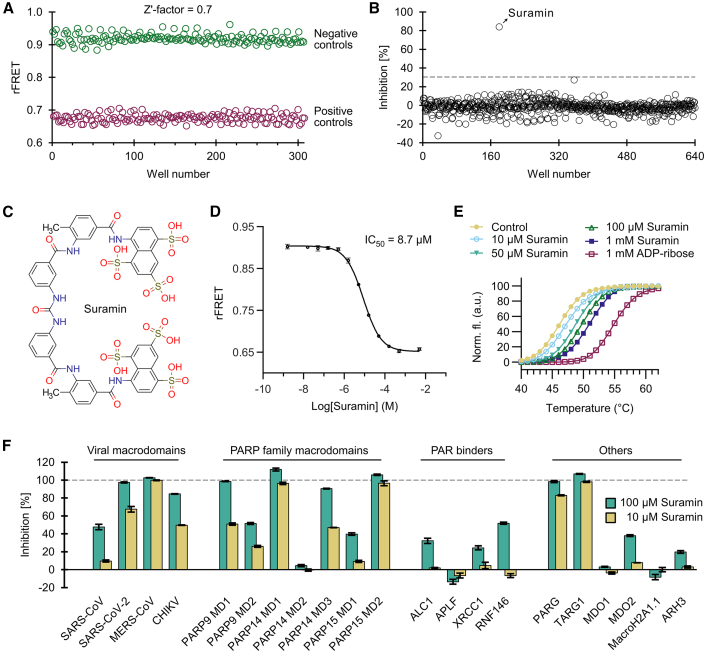
Proteins interacting with ADP-ribosyl groups are often involved in disease-related pathways or viral infections, making them attractive drug targets. We present a robust and accessible assay applicable to both hydrolyzing or non-hydrolyzing binders of mono- and poly-ADP-ribosyl groups. This technology relies on a C-terminal tag based on a Gi protein alpha subunit peptide (GAP), which allows for site-specific introduction of cysteine-linked mono- and poly-ADP-ribosyl groups or analogs. By fusing the GAP-tag and ADP-ribosyl binders to fluorescent proteins, we generate robust FRET partners and confirm the interaction with 22 known ADP-ribosyl binders. The applicability for high-throughput screening of inhibitors is demonstrated with the SARS-CoV-2 nsp3 macrodomain, for which we identify suramin as a moderate-affinity yet non-specific inhibitor. High-affinity ADP-ribosyl binders fused to nanoluciferase complement this technology, enabling simple blot-based detection of ADP-ribosylated proteins. All these tools can be produced in Escherichia coli and will help in ADP-ribosylation research and drug discovery.
Keywords: ADP-ribosylation; SARS-CoV-2; binding assay; high-throughput screening; inhibitors; macrodomain; post-translational modification; protein labeling.
© 2021 The Author(s).
Conflict of interest statement
S.T.S., A.G.P., and L.L. are inventors listed in a patent application related to the described methods, and these authors declare no additional interests. The remaining authors declare no competing interests.
Figures
Similar articles
-
The SARS-CoV-2 Nsp3 macrodomain reverses PARP9/DTX3L-dependent ADP-ribosylation induced by interferon signaling.J Biol Chem. 2021 Sep;297(3):101041. doi: 10.1016/j.jbc.2021.101041. Epub 2021 Aug 4. J Biol Chem. 2021. PMID: 34358560 Free PMC article.
-
Development of an Inhibitor Screening Assay for Mono-ADP-Ribosyl Hydrolyzing Macrodomains Using AlphaScreen Technology.SLAS Discov. 2018 Mar;23(3):255-263. doi: 10.1177/2472555217737006. Epub 2017 Oct 13. SLAS Discov. 2018. PMID: 29028410
-
Host ADP-ribosylation and the SARS-CoV-2 macrodomain.Biochem Soc Trans. 2021 Aug 27;49(4):1711-1721. doi: 10.1042/BST20201212. Biochem Soc Trans. 2021. PMID: 34351418 Free PMC article. Review.
-
Activity-Based Screening Assay for Mono-ADP-Ribosylhydrolases.SLAS Discov. 2021 Jan;26(1):67-76. doi: 10.1177/2472555220928911. Epub 2020 Jun 12. SLAS Discov. 2021. PMID: 32527186
-
ADP-ribosylhydrolases: from DNA damage repair to COVID-19.J Zhejiang Univ Sci B. 2021 Jan 15;22(1):21-30. doi: 10.1631/jzus.B2000319. J Zhejiang Univ Sci B. 2021. PMID: 33448184 Free PMC article. Review.
Cited by
-
High-throughput screening assay for PARP-HPF1 interaction inhibitors to affect DNA damage repair.Sci Rep. 2024 Feb 16;14(1):3875. doi: 10.1038/s41598-024-54123-8. Sci Rep. 2024. PMID: 38365924 Free PMC article.
-
Intracellular mono-ADP-ribosyltransferases at the host-virus interphase.Cell Mol Life Sci. 2022 May 10;79(6):288. doi: 10.1007/s00018-022-04290-6. Cell Mol Life Sci. 2022. PMID: 35536484 Free PMC article. Review.
-
SARS-CoV-2 Mac1 is required for IFN antagonism and efficient virus replication in mice.bioRxiv [Preprint]. 2023 Apr 6:2023.04.06.535927. doi: 10.1101/2023.04.06.535927. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2023 Aug 29;120(35):e2302083120. doi: 10.1073/pnas.2302083120. PMID: 37066301 Free PMC article. Updated. Preprint.
-
The zinc-binding motif in tankyrases is required for the structural integrity of the catalytic ADP-ribosyltransferase domain.Open Biol. 2022 Mar;12(3):210365. doi: 10.1098/rsob.210365. Epub 2022 Mar 23. Open Biol. 2022. PMID: 35317661 Free PMC article.
-
An Update on the Current State of SARS-CoV-2 Mac1 Inhibitors.Pathogens. 2023 Oct 7;12(10):1221. doi: 10.3390/pathogens12101221. Pathogens. 2023. PMID: 37887737 Free PMC article. Review.
References
-
- Abraham R., Hauer D., McPherson R.L., Utt A., Kirby I.T., Cohen M.S., Merits A., Leung A.K.L., Griffin D.E. ADP-ribosyl-binding and hydrolase activities of the alphavirus nsP3 macrodomain are critical for initiation of virus replication. Proc. Natl. Acad. Sci. U S A. 2018;115:E10457–E10466. - PMC - PubMed
-
- Ahel I., Ahel D., Matsusaka T., Clark A.J., Pines J., Boulton S.J., West S.C. Poly(ADP-ribose)-binding zinc finger motifs in DNA repair/checkpoint proteins. Nature. 2008;451:81–85. - PubMed
-
- Albulescu I.C., van Hoolwerff M., Wolters L.A., Bottaro E., Nastruzzi C., Yang S.C., Tsay S.-C., Hwu J.R., Snijder E.J., van Hemert M.J. Suramin inhibits chikungunya virus replication through multiple mechanisms. Antivir. Res. 2015;121:39–46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
