

Deleting DNMT3A in CAR T cells prevents exhaustion and enhances antitumor activity
- PMID: 34788079
- PMCID: PMC8733956
- DOI: 10.1126/scitranslmed.abh0272
Deleting DNMT3A in CAR T cells prevents exhaustion and enhances antitumor activity
Abstract
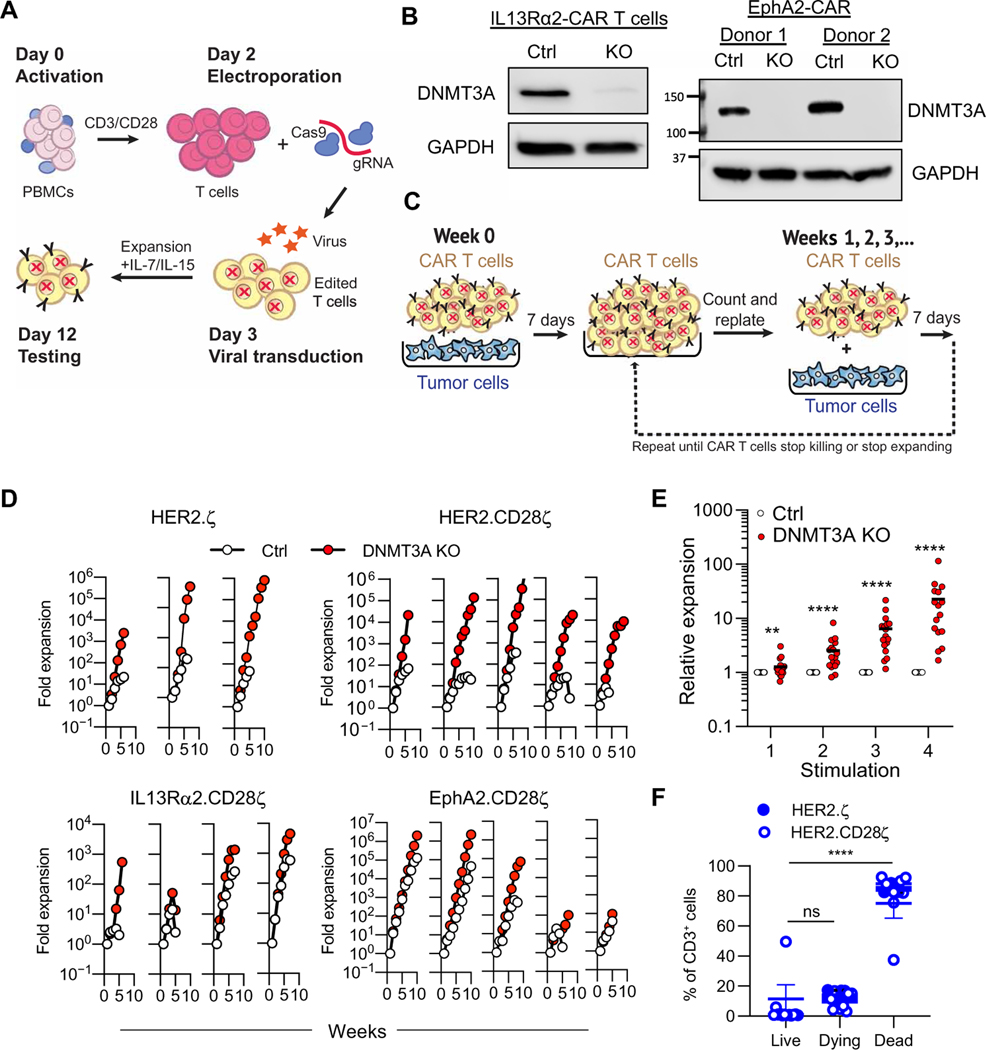
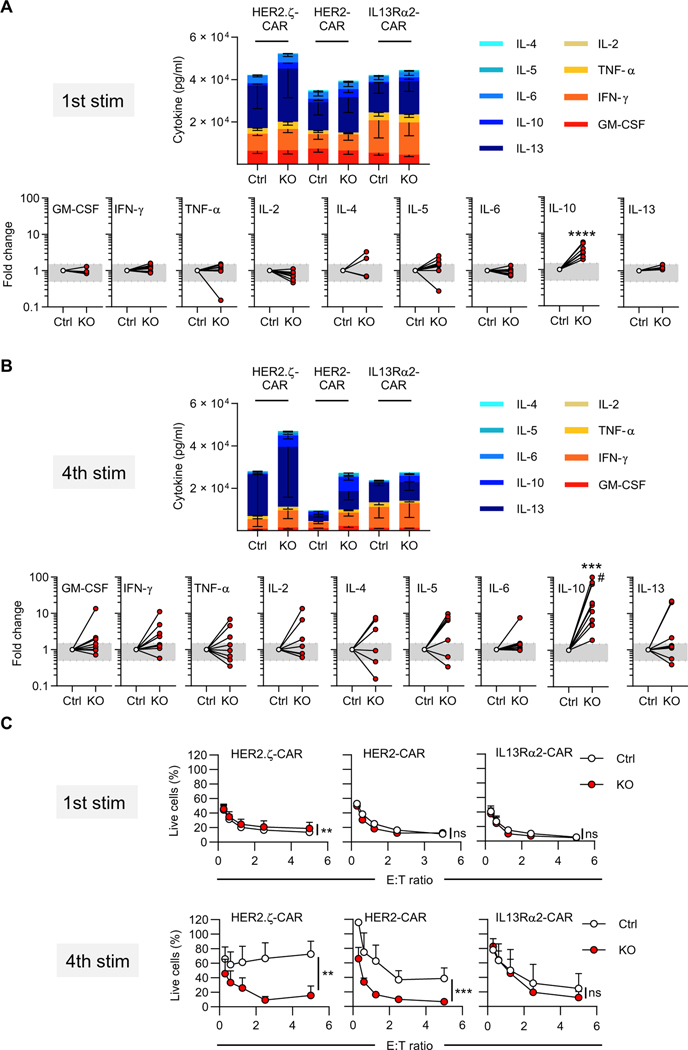
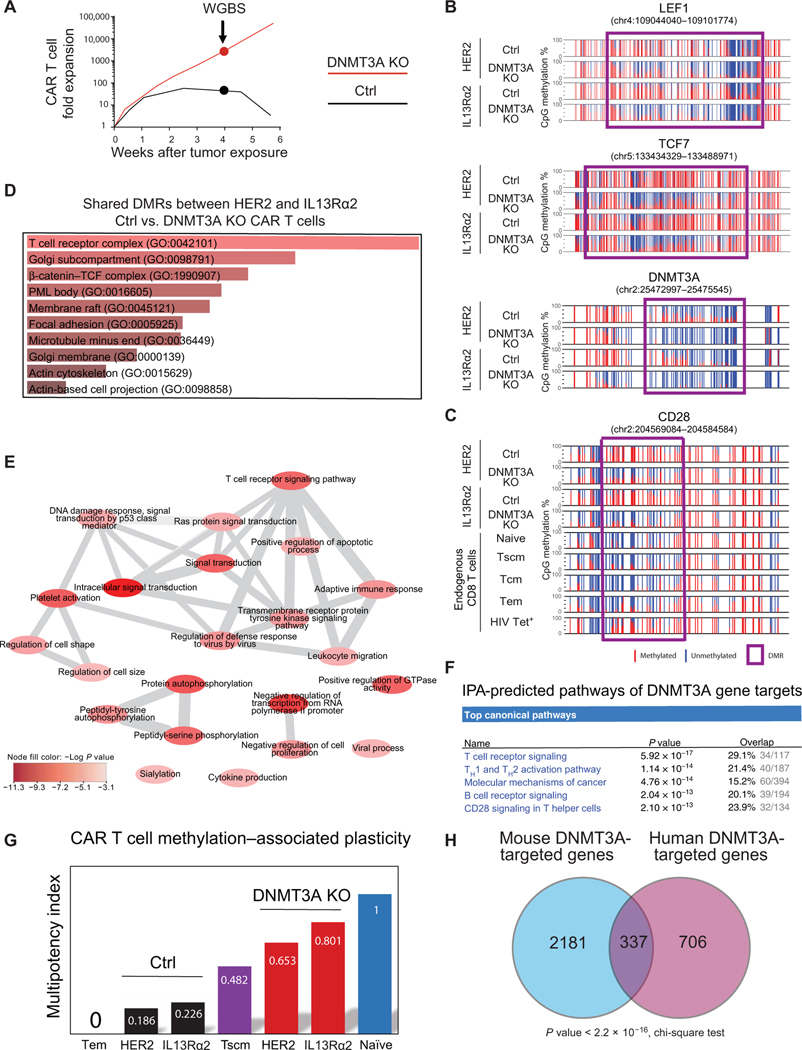
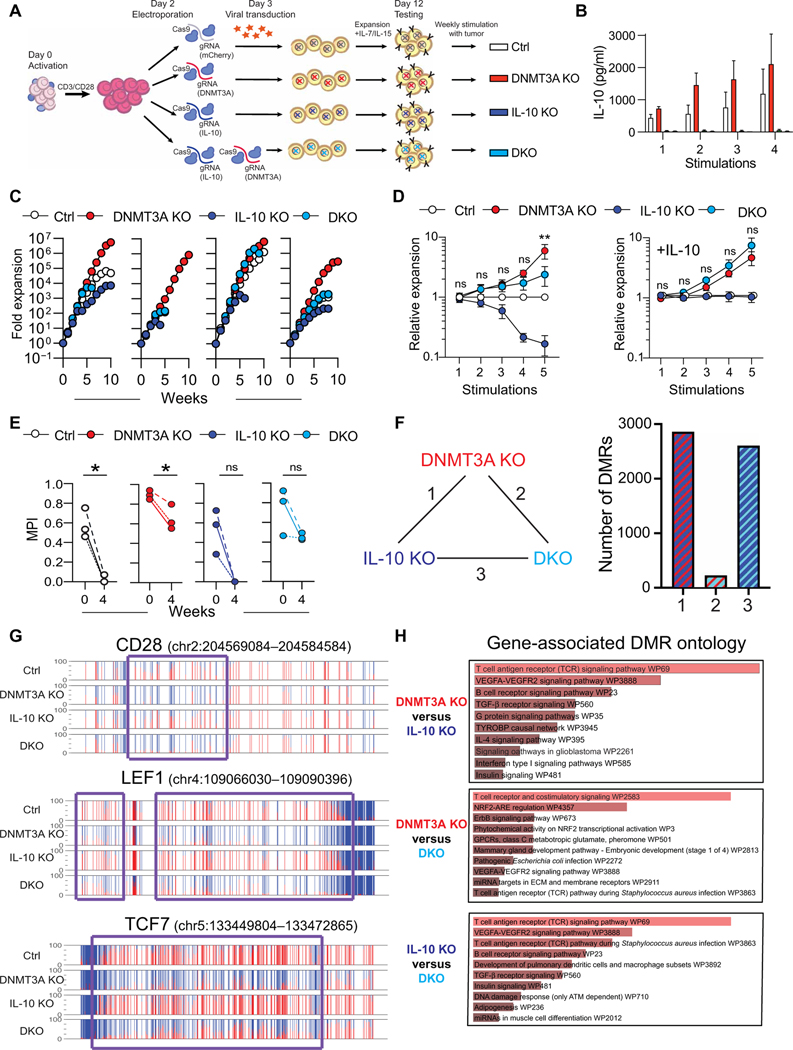
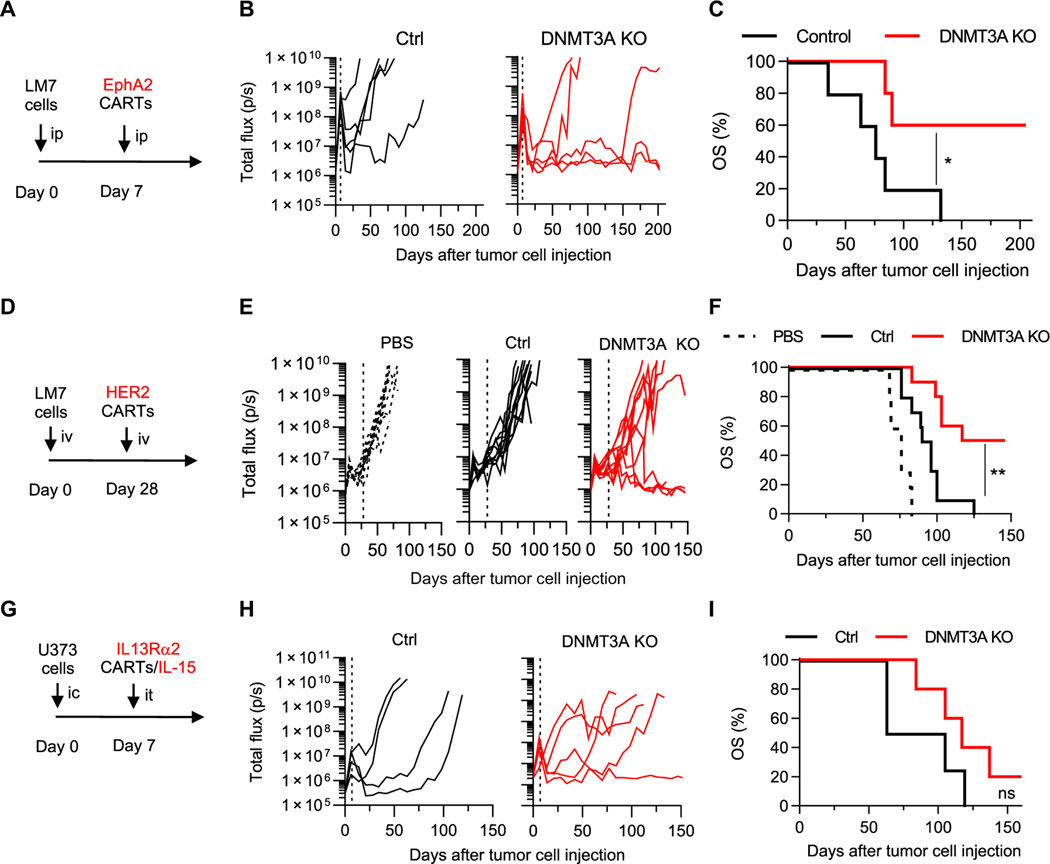
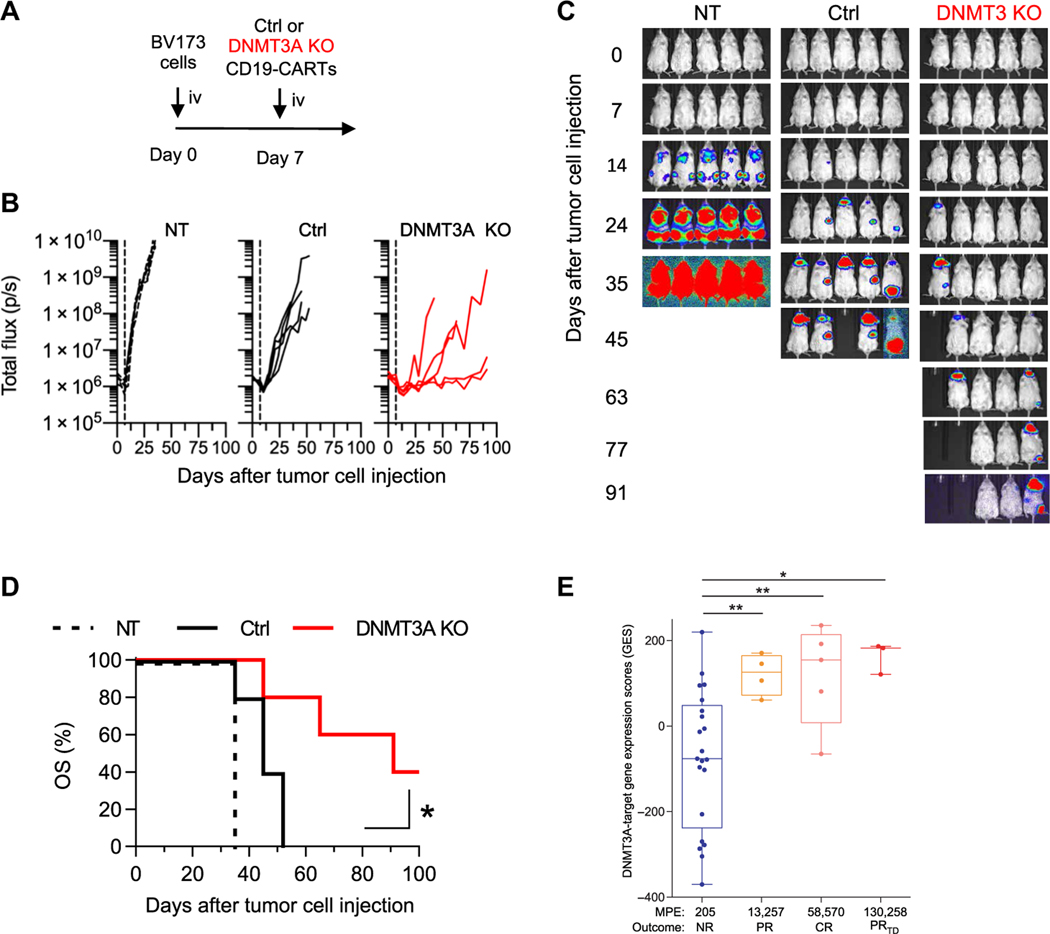
Chimeric antigen receptor (CAR) T cell therapy is revolutionizing cancer immunotherapy for patients with B cell malignancies and is now being developed for solid tumors and chronic viral infections. Although clinical trials have demonstrated the curative potential of CAR T cell therapy, a substantial and well-established limitation is the heightened contraction and transient persistence of CAR T cells during prolonged antigen exposure. The underlying mechanism(s) for this dysfunctional state, often termed CAR T cell exhaustion, remains poorly defined. Here, we report that exhaustion of human CAR T cells occurs through an epigenetic repression of the T cell’s multipotent developmental potential. Deletion of the de novo DNA methyltransferase 3 alpha (DNMT3A) in T cells expressing first- or second-generation CARs universally preserved the cells’ ability to proliferate and mount an antitumor response during prolonged tumor exposure. The increased functionality of the exhaustion-resistant DNMT3A knockout CAR T cells was coupled to an up-regulation of interleukin-10, and genome-wide DNA methylation profiling defined an atlas of genes targeted for epigenetic silencing. This atlas provides a molecular definition of CAR T cell exhaustion, which includes many transcriptional regulators that limit the “stemness” of immune cells, including CD28, CCR7, TCF7, and LEF1. Last, we demonstrate that this epigenetically regulated multipotency program is firmly coupled to the clinical outcome of prior CAR T cell therapies. These data document the critical role epigenetic mechanisms play in limiting the fate potential of human T cells and provide a road map for leveraging this information for improving CAR T cell efficacy.
Figures
References
-
- Maude SL, Laetsch TW, Buechner J, Rives S, Boyer M, Bittencourt H, Bader P, Verneris MR, Stefanski HE, Myers GD, Qayed M, De Moerloose B, Hiramatsu H, Schlis K, Davis KL, Martin PL, Nemecek ER, Yanik GA, Peters C, Baruchel A, Boissel N, Mechinaud F, Balduzzi A, Krueger J, June CH, Levine BL, Wood P, Taran T, Leung M, Mueller KT, Zhang Y, Sen K, Lebwohl D, Pulsipher MA, Grupp SA, Tisagenlecleucel in children and young adults with B-cell lymphoblastic leukemia. N. Engl. J. Med. 378, 439–448 (2018). - PMC - PubMed
-
- Fraietta JA, Lacey SF, Orlando EJ, Pruteanu-Malinici I, Gohil M, Lundh S, Boesteanu AC, Wang Y, O’Connor RS, Hwang WT, Pequignot E, Ambrose DE, Zhang C, Wilcox N, Bedoya F, Dorfmeier C, Chen F, Tian L, Parakandi H, Gupta M, Young RM, Johnson FB, Kulikovskaya I, Liu L, Xu J, Kassim SH, Davis MM, Levine BL, Frey NV, Siegel DL, Huang AC, Wherry EJ, Bitter H, Brogdon JL, Porter DL, June CH, Melenhorst JJ, Determinants of response and resistance to CD19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat. Med. 24, 563–571 (2018). - PMC - PubMed
-
- Ahmed N, Brawley VS, Hegde M, Robertson C, Ghazi A, Gerken C, Liu E, Dakhova O, Ashoori A, Corder A, Gray T, Wu MF, Liu H, Hicks J, Rainusso N, Dotti G, Mei Z, Grilley B, Gee A, Rooney CM, Brenner MK, Heslop HE, Wels WS, Wang LL, Anderson P, Gottschalk S, Human epidermal growth factor receptor 2 (HER2)-specific chimeric antigen receptor-modified T cells for the immunotherapy of HER2-positive sarcoma. J. Clin. Oncol. 33, 1688–1696 (2015). - PMC - PubMed
-
- Beatty GL, O’Hara MH, Lacey SF, Torigian DA, Nazimuddin F, Chen F, Kulikovskaya IM, Soulen MC, McGarvey M, Nelson AM, Gladney WL, Levine BL, Melenhorst JJ, Plesa G, June CH, Activity of mesothelin-specific chimeric antigen receptor t cells against pancreatic carcinoma metastases in a phase 1 trial. Gastroenterology 155, 29–32 (2018). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K99 CA256262/CA/NCI NIH HHS/United States
- UM1 AI109565/AI/NIAID NIH HHS/United States
- R01 CA173750/CA/NCI NIH HHS/United States
- F31 CA250401/CA/NCI NIH HHS/United States
- R01 CA237311/CA/NCI NIH HHS/United States
- R01 NS106379/NS/NINDS NIH HHS/United States
- P30 CA021765/CA/NCI NIH HHS/United States
- P01 CA096832/CA/NCI NIH HHS/United States
- R01 NS121249/NS/NINDS NIH HHS/United States
- R50 CA211481/CA/NCI NIH HHS/United States
- R01 AI114442/AI/NIAID NIH HHS/United States
- U01 AI157189/AI/NIAID NIH HHS/United States
- R00 CA256262/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
