

Phylogenomic approaches to detecting and characterizing introgression
- PMID: 34788444
- PMCID: PMC9208645
- DOI: 10.1093/genetics/iyab173
Phylogenomic approaches to detecting and characterizing introgression
Erratum in
-
Corrigendum to: Phylogenomic approaches to detecting and characterizing introgression.Genetics. 2022 Feb 4;220(2):iyab220. doi: 10.1093/genetics/iyab220. Genetics. 2022. PMID: 35100344 Free PMC article. No abstract available.
Abstract
Phylogenomics has revealed the remarkable frequency with which introgression occurs across the tree of life. These discoveries have been enabled by the rapid growth of methods designed to detect and characterize introgression from whole-genome sequencing data. A large class of phylogenomic methods makes use of data across species to infer and characterize introgression based on expectations from the multispecies coalescent. These methods range from simple tests, such as the D-statistic, to model-based approaches for inferring phylogenetic networks. Here, we provide a detailed overview of the various signals that different modes of introgression are expected leave in the genome, and how current methods are designed to detect them. We discuss the strengths and pitfalls of these approaches and identify areas for future development, highlighting the different signals of introgression, and the power of each method to detect them. We conclude with a discussion of current challenges in inferring introgression and how they could potentially be addressed.
Keywords: hybridization; introgression; phylogenomic methods; phylogenomics.
© The Author(s) 2021. Published by Oxford University Press on behalf of Genetics Society of America.
Figures
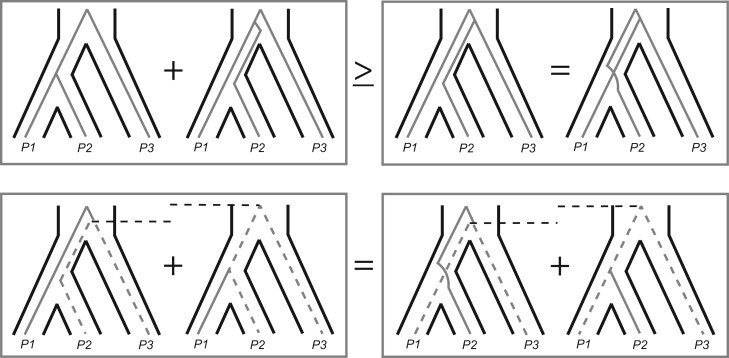
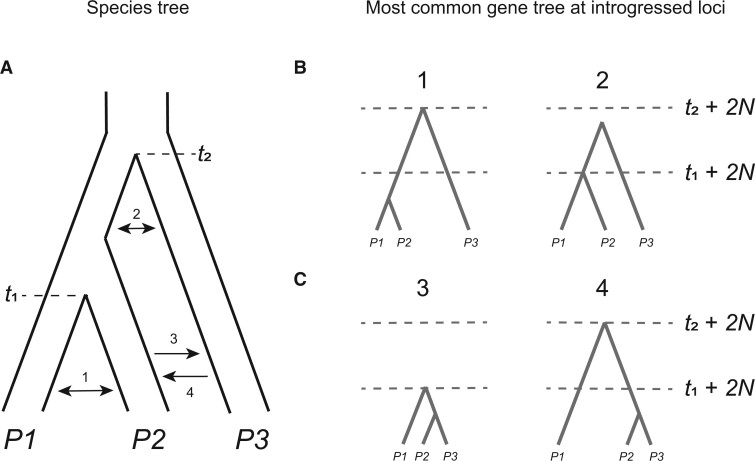
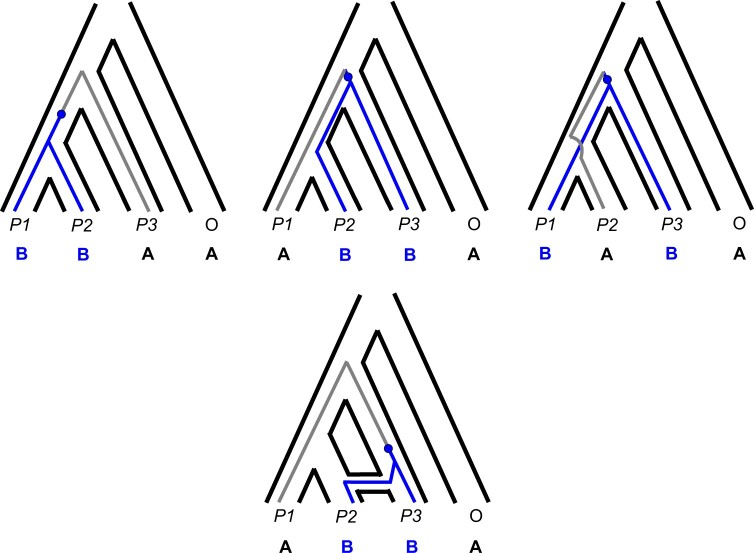
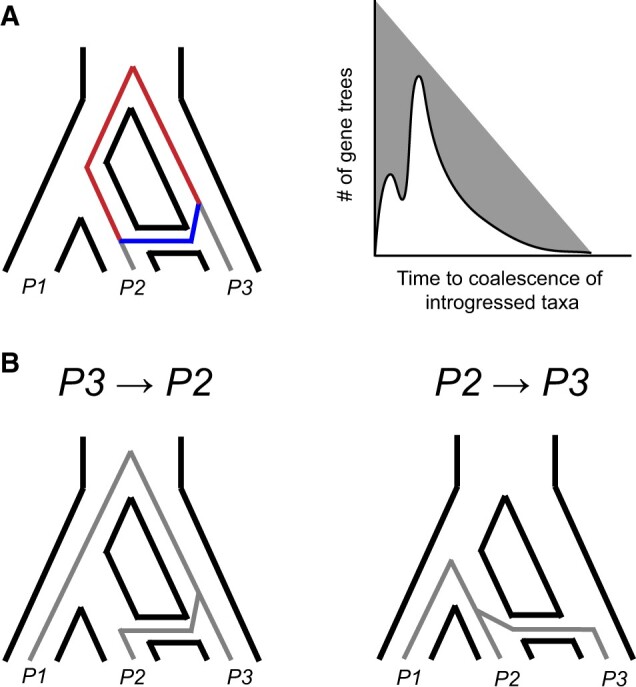
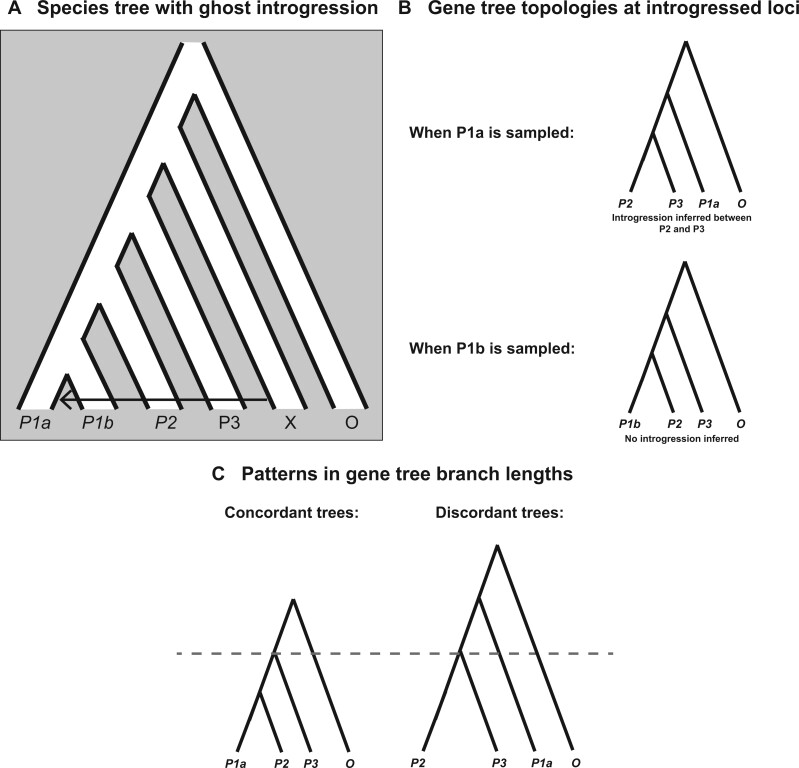
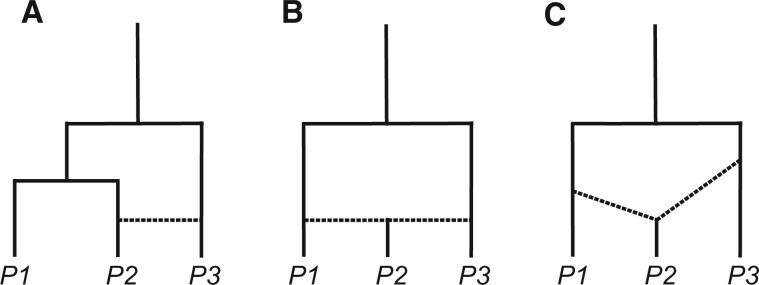
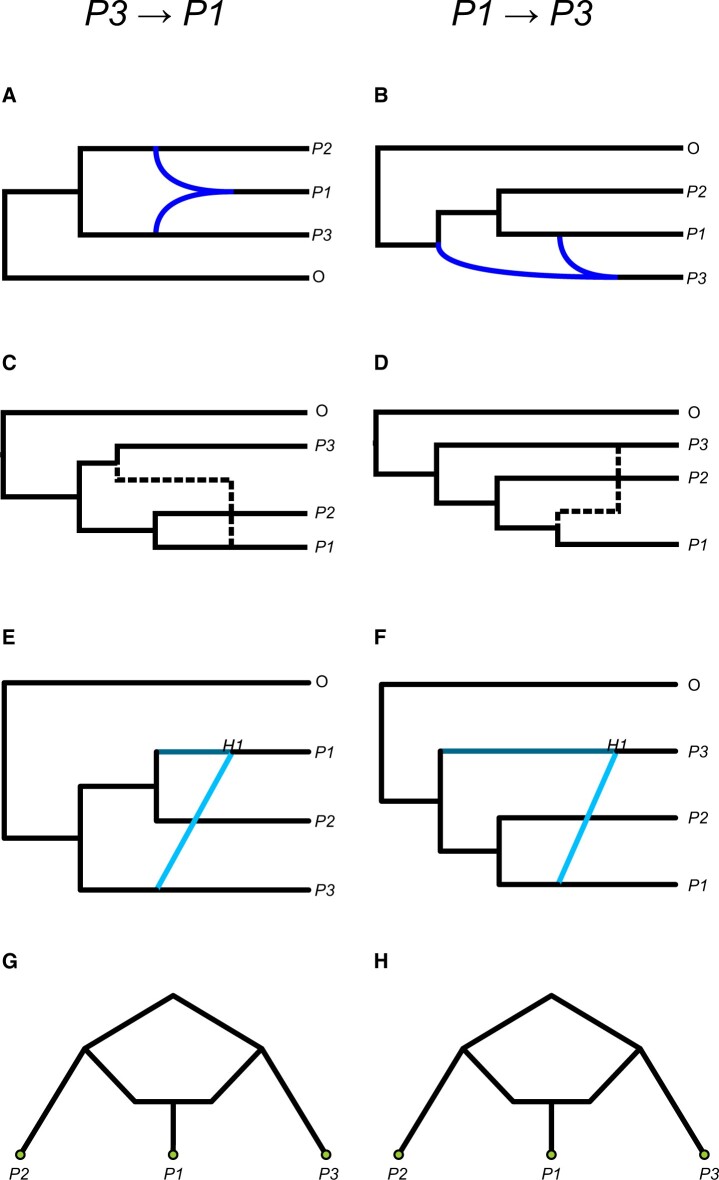
References
-
- Adams RH, Schield DR, Card DC, Castoe TA.. 2018. Assessing the impacts of positive selection on coalescent-based species tree estimation and species delimitation. Syst Biol. 67:1076–1090. - PubMed
-
- Ai H, Fang X, Yang B, Huang Z, Chen H, et al.2015. Adaptation and possible ancient interspecies introgression in pigs identified by whole-genome sequencing. Nat Genet. 47:217–225. - PubMed
-
- Akaike H. 1974. A new look at the statistical model identification. IEEE Trans Automat Contr. 19:716–723.
-
- Baum DA. 2007. Concordance trees, concordance factors, and the exploration of reticulate genealogy. Taxon. 56:417–426.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
