

Innate Immunity and Cancer Pathophysiology
- PMID: 34788549
- PMCID: PMC9012188
- DOI: 10.1146/annurev-pathmechdis-032221-115501
Innate Immunity and Cancer Pathophysiology
Abstract
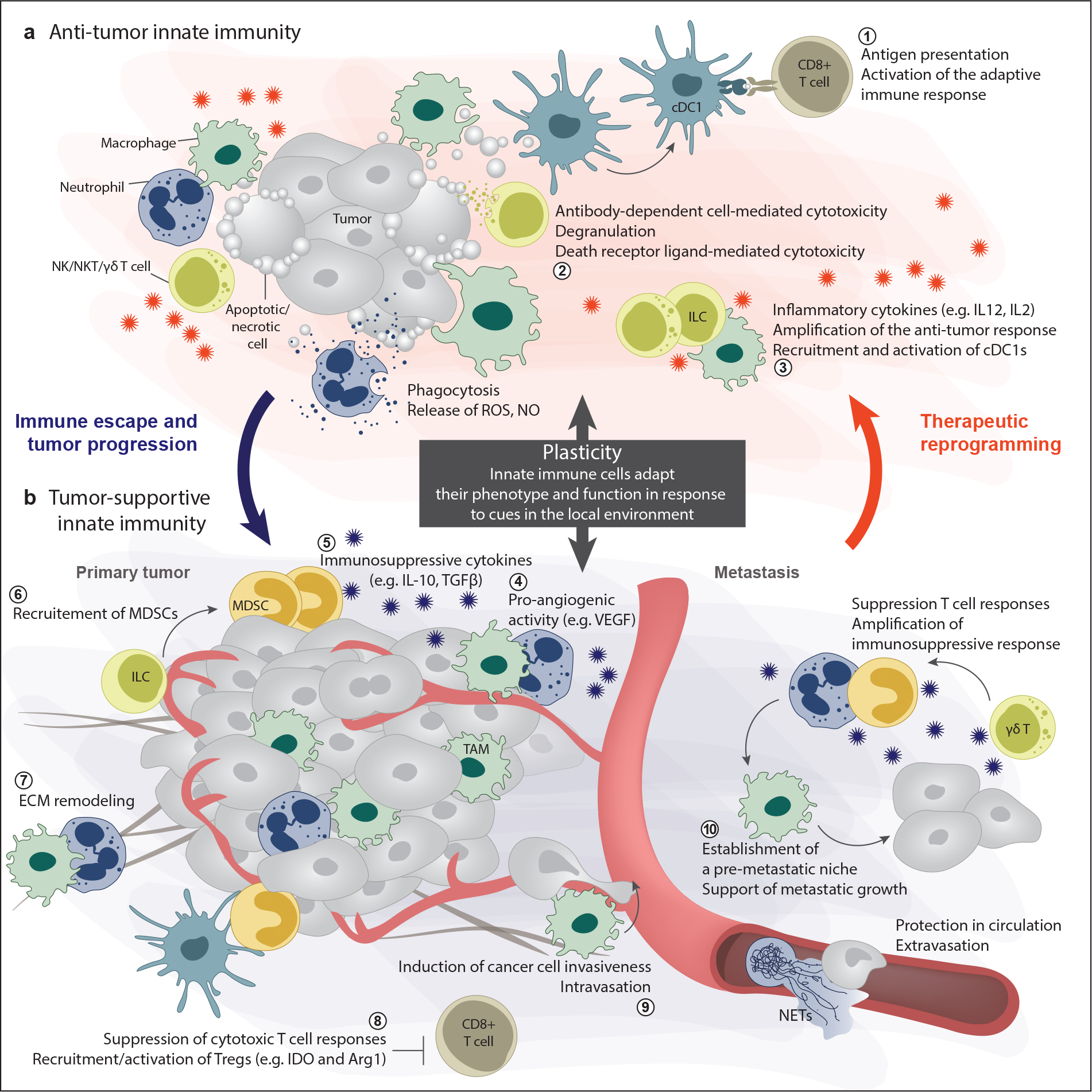
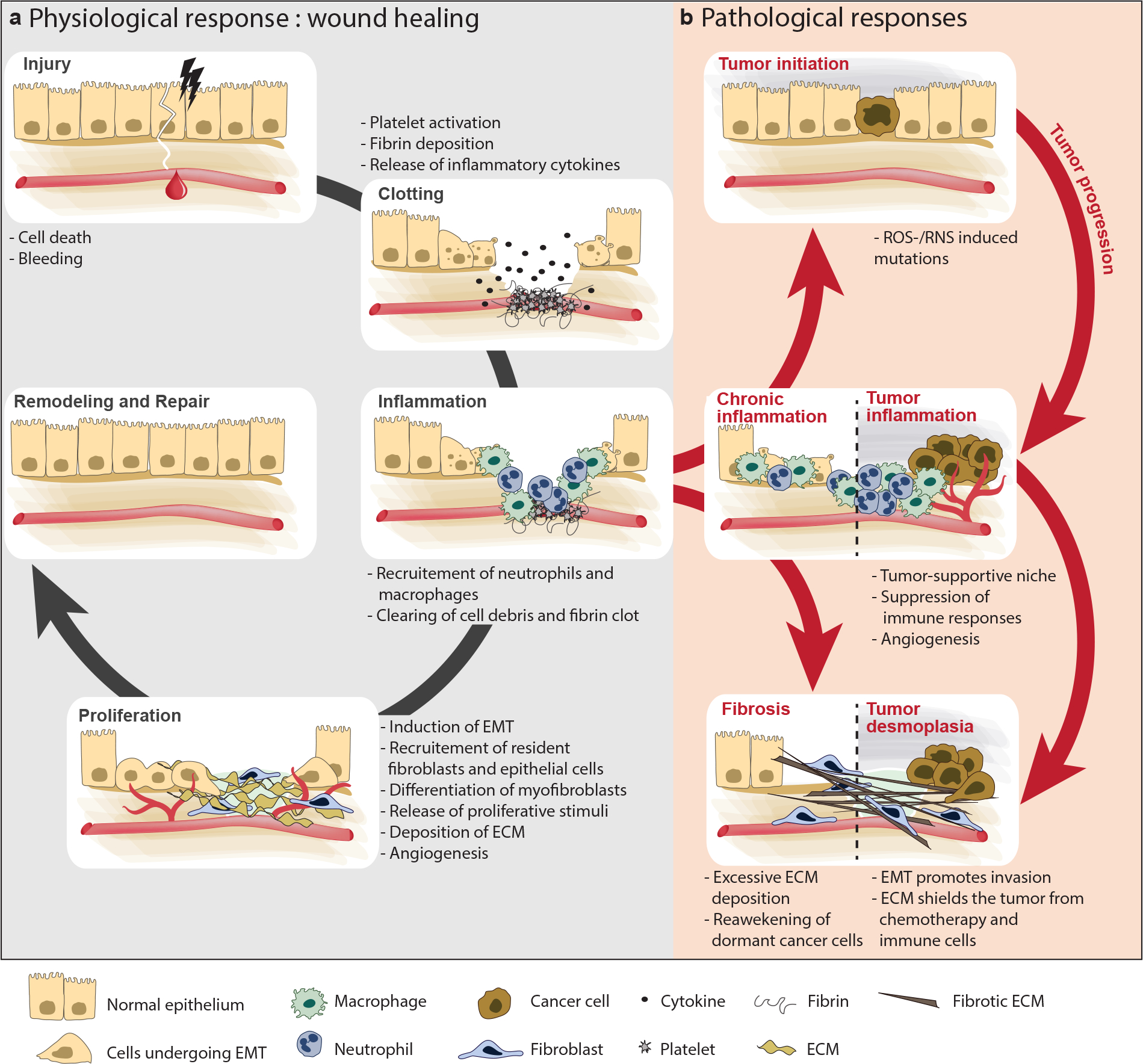
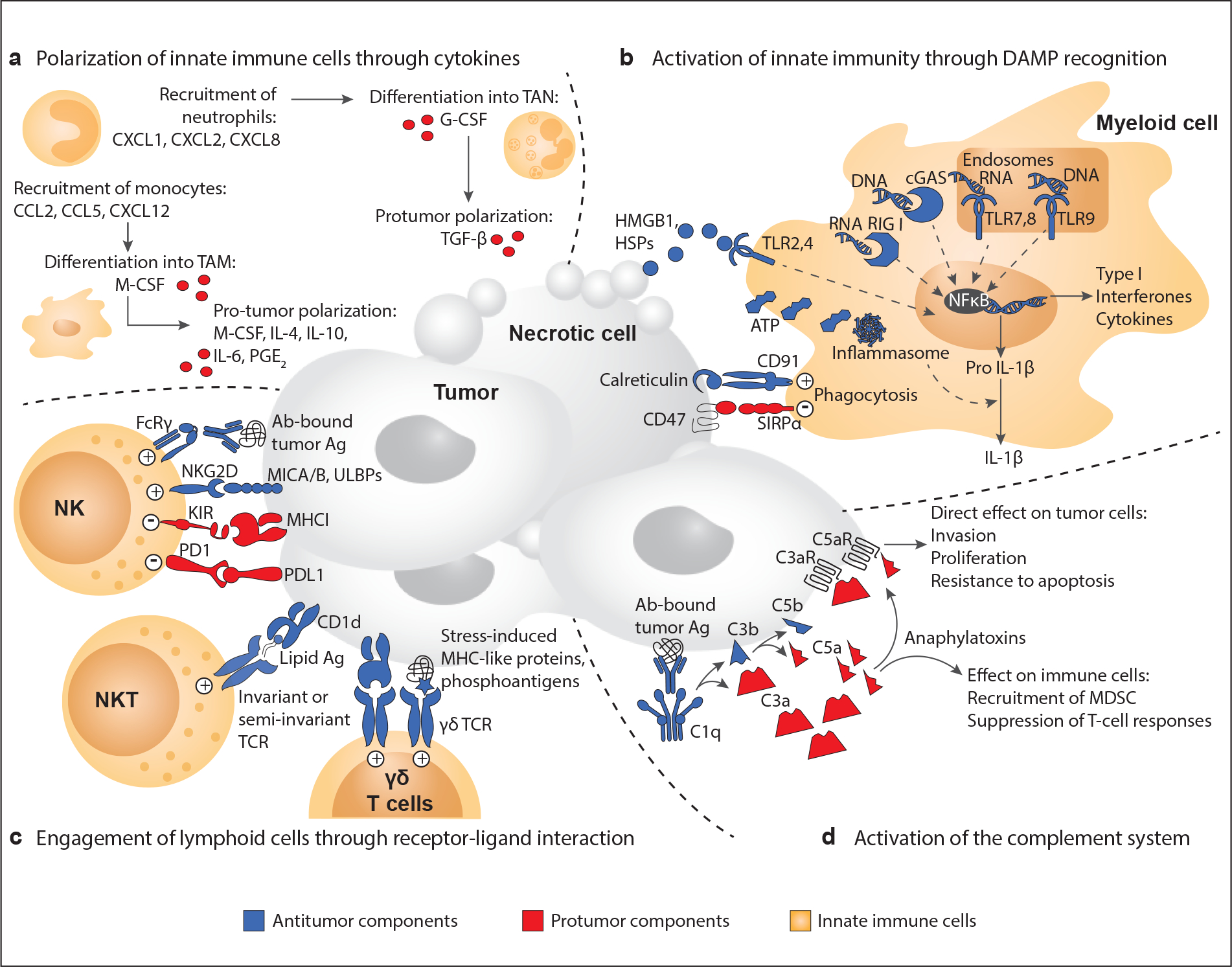
Chronic inflammation increases the risk of several cancers, including gastric, colon, and hepatic cancers. Conversely, tumors, similar to tissue injury, trigger an inflammatory response coordinated by the innate immune system. Cellular and molecular mediators of inflammation modulate tumor growth directly and by influencing the adaptive immune response. Depending on the balance of immune cell types and signals within the tumor microenvironment, inflammation can support or restrain the tumor. Adding to the complexity, research from the past two decades has revealed that innate immune cells are highly heterogeneous and plastic, with variable phenotypes depending on tumor type, stage, and treatment. The field is now on the cusp of being able to harness this wealth of data to (a) classify tumors on the basis of their immune makeup, with implications for prognosis, treatment choice, and clinical outcome, and (b) design therapeutic strategies that activate antitumor immune responses by targeting innate immune cells.
Keywords: cancer immunology; immuno-oncology; inflammation; innate immunity; plasticity; tumor microenvironment.
Figures
References
-
- Virchow R 1989. (1858). Cellular pathology. As based upon physiological and pathological histology. Lecture XVI--Atheromatous affection of arteries. 1858. Nutr. Rev. 47:23–25 - PubMed
-
- Coley WB. 1991. The treatment of malignant tumors by repeated inoculations of erysipelas. With a report of ten original cases. 1893. Clin Orthop Relat Res: 3–11 - PubMed
-
- Starnes CO. 1992. Coley’s toxins in perspective. Nature 357: 11–2 - PubMed
-
- Balkwill F, Mantovani A. 2001. Inflammation and cancer: back to Virchow? Lancet 357: 539–45 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
