

A NYN domain protein directly interacts with DECAPPING1 and is required for phyllotactic pattern
- PMID: 34791434
- PMCID: PMC8825452
- DOI: 10.1093/plphys/kiab529
A NYN domain protein directly interacts with DECAPPING1 and is required for phyllotactic pattern
Abstract
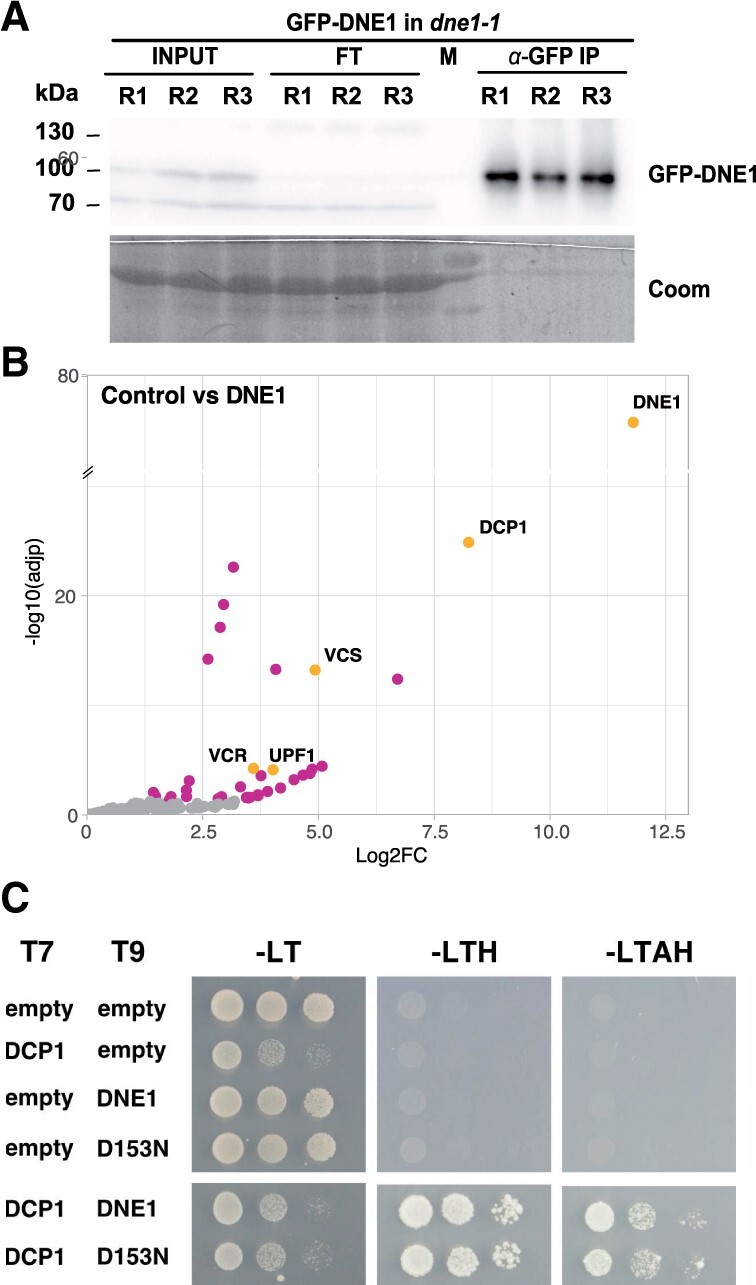
In eukaryotes, general mRNA decay requires the decapping complex. The activity of this complex depends on its catalytic subunit, DECAPPING2 (DCP2), and its interaction with decapping enhancers, including its main partner DECAPPING1 (DCP1). Here, we report that in Arabidopsis thaliana, DCP1 also interacts with a NYN domain endoribonuclease, hence named DCP1-ASSOCIATED NYN ENDORIBONUCLEASE 1 (DNE1). Interestingly, we found DNE1 predominantly associated with DCP1, but not with DCP2, and reciprocally, suggesting the existence of two distinct protein complexes. We also showed that the catalytic residues of DNE1 are required to repress the expression of mRNAs in planta upon transient expression. The overexpression of DNE1 in transgenic lines led to growth defects and a similar gene deregulation signature than inactivation of the decapping complex. Finally, the combination of dne1 and dcp2 mutations revealed a functional redundancy between DNE1 and DCP2 in controlling phyllotactic pattern formation. Our work identifies DNE1, a hitherto unknown DCP1 protein partner highly conserved in the plant kingdom and identifies its importance for developmental robustness.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures







References
-
- Anderson SJ, Kramer MC, Gosai SJ, Yu X, Vandivier LE, Nelson ADL, Anderson ZD, Beilstein MA, Fray RG, Lyons E, et al. (2018) N6-methyladenosine inhibits local ribonucleolytic cleavage to stabilize mRNAs in Arabidopsis. Cell Rep 25: 1146–1157.e3 - PubMed
-
- Bassal M, Abukhalaf M, Majovsky P, Thieme D, Herr T, Ayash M, Tabassum N, Al Shweiki MR, Proksch C, Hmedat A, et al. (2020) Reshaping of the Arabidopsis thaliana proteome landscape and co-regulation of proteins in development and immunity. Mol Plant 13: 1709–1732 - PubMed
-
- Besnard F, Refahi Y, Morin V, Marteaux B, Brunoud G, Chambrier P, Rozier F, Mirabet V, Legrand J, Lainé S, et al. (2014) Cytokinin signalling inhibitory fields provide robustness to phyllotaxis. Nature 505: 417–421 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
