

Interspecies transcriptomics identify genes that underlie disproportionate foot growth in jerboas
- PMID: 34793695
- PMCID: PMC8792248
- DOI: 10.1016/j.cub.2021.10.063
Interspecies transcriptomics identify genes that underlie disproportionate foot growth in jerboas
Abstract
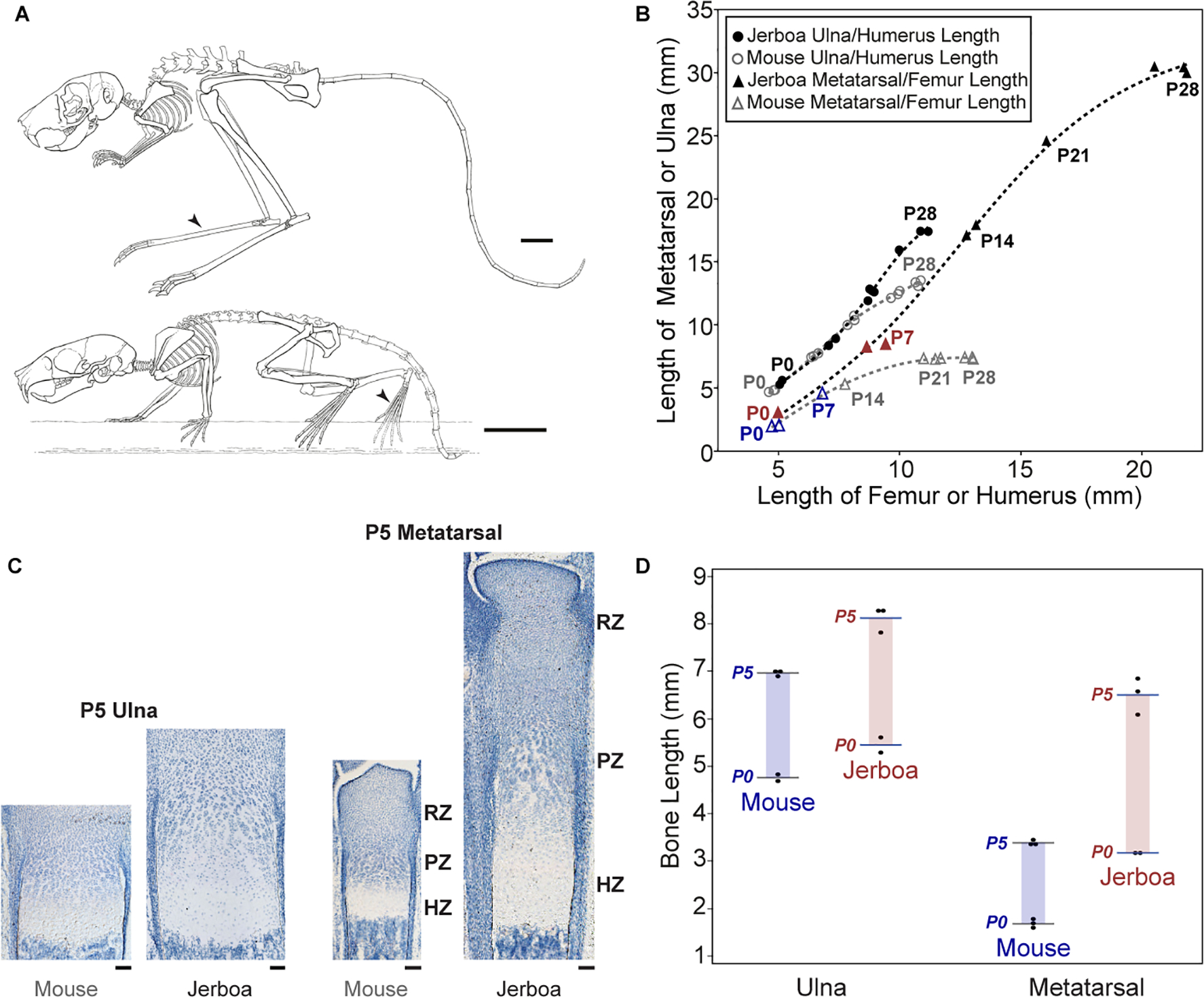
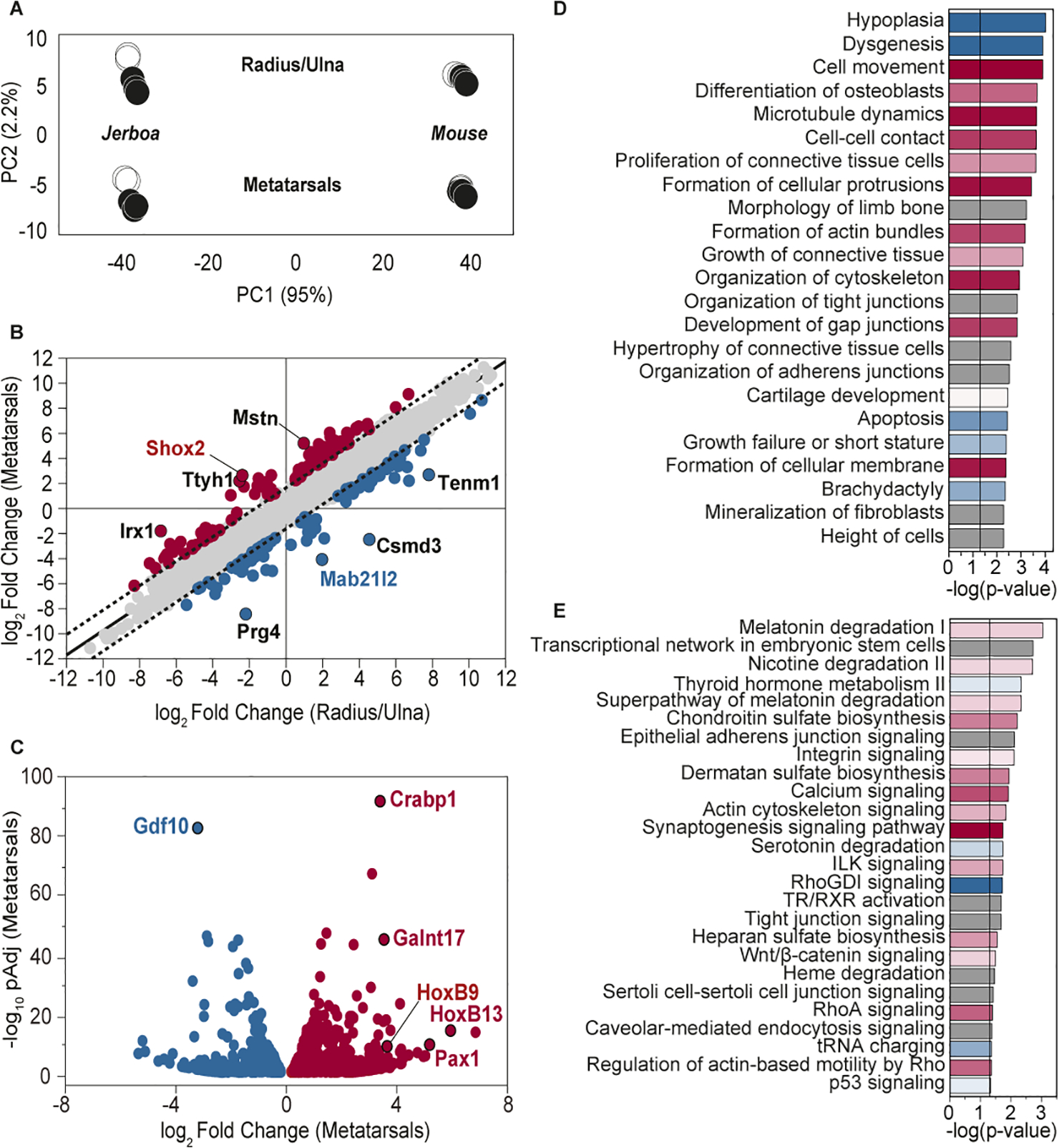
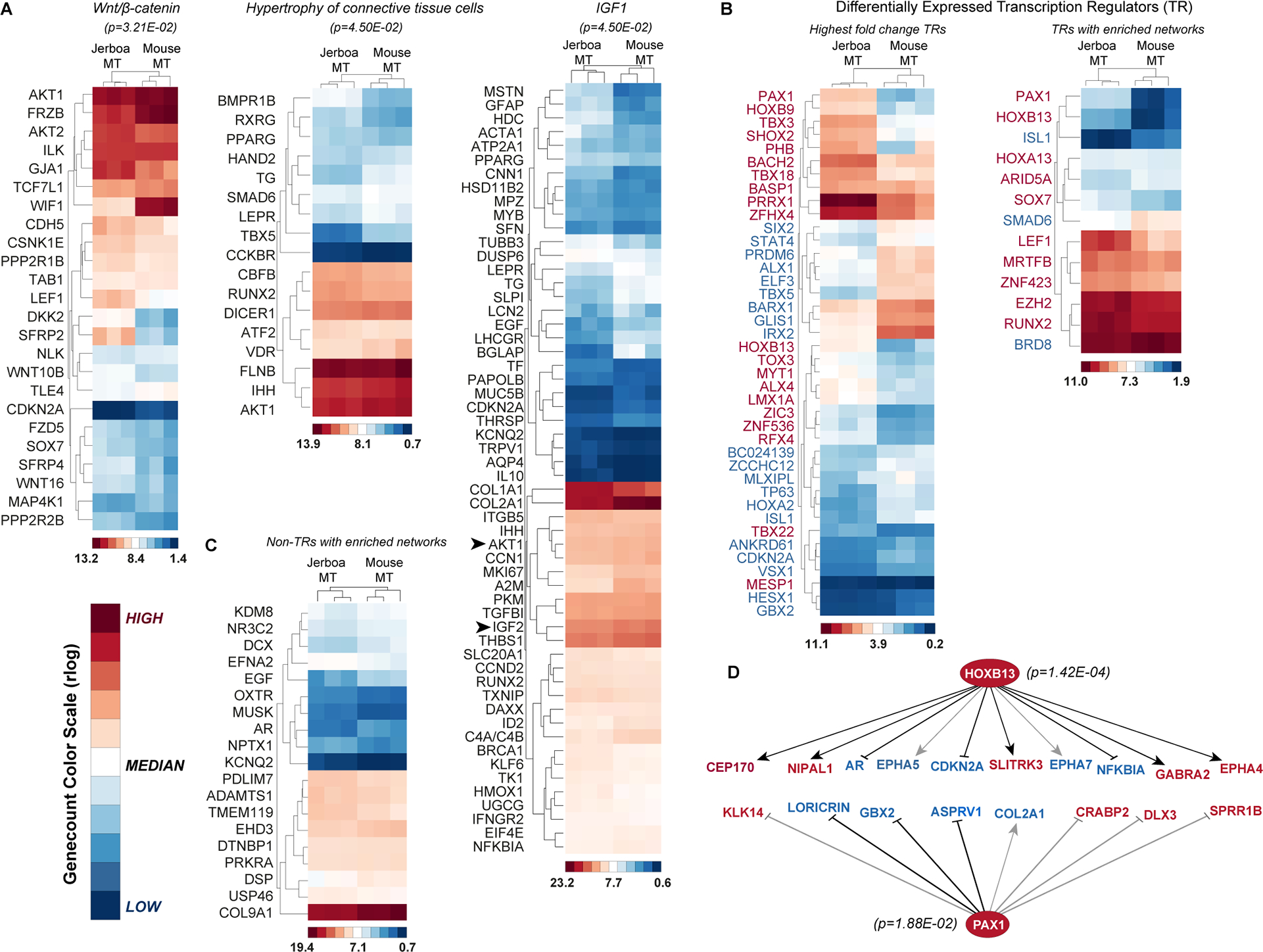
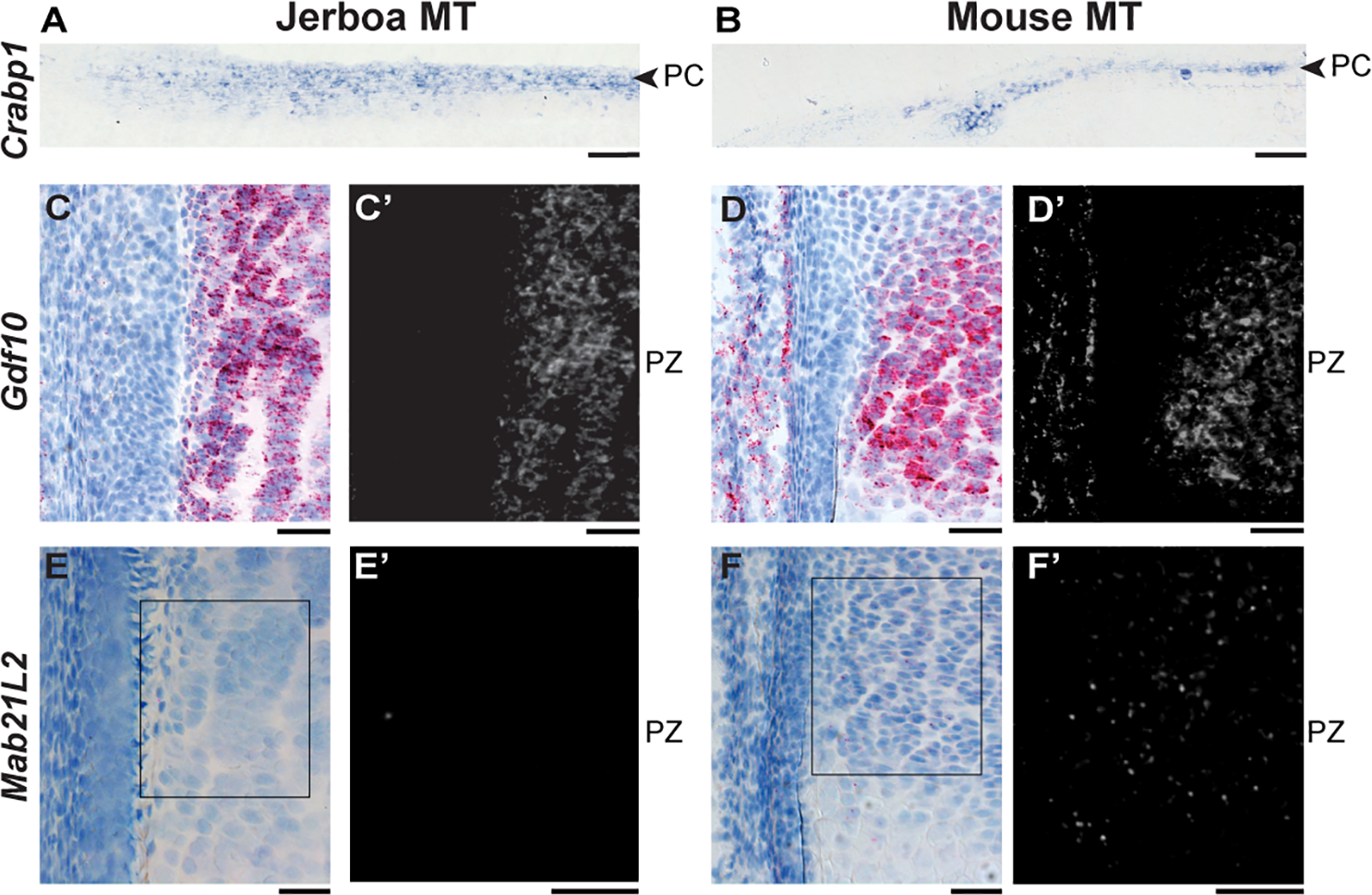
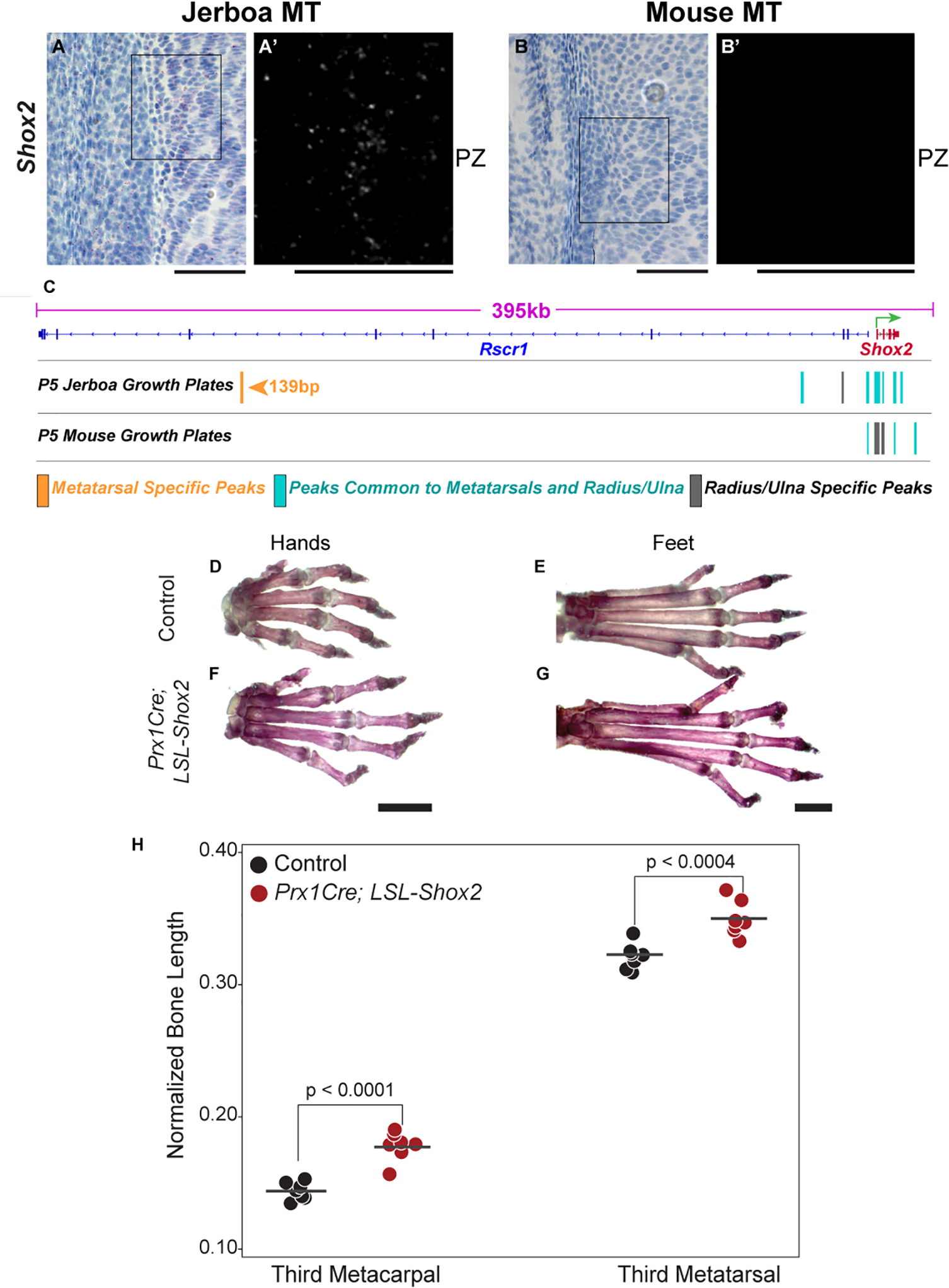
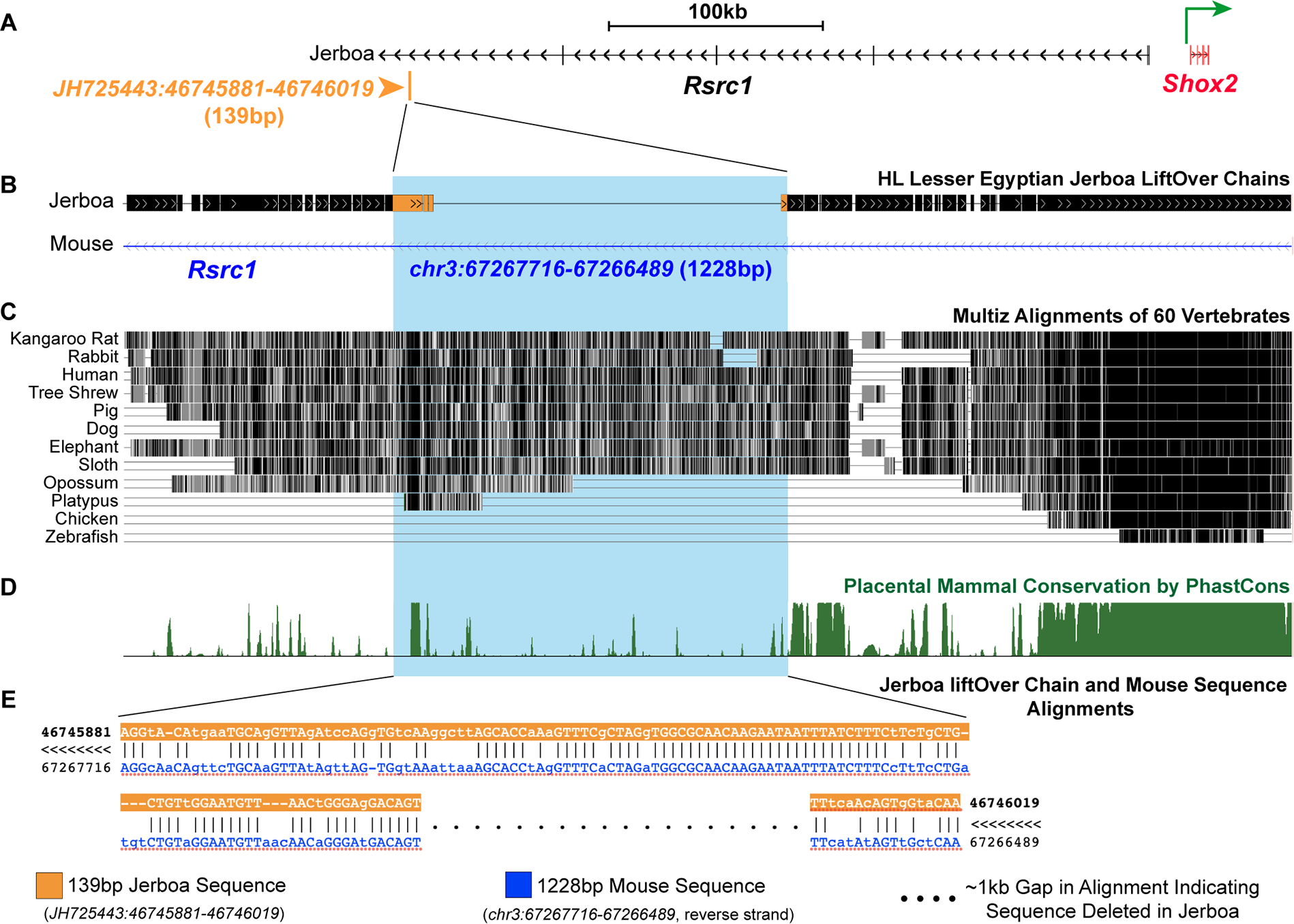
Despite the great diversity of vertebrate limb proportion and our deep understanding of the genetic mechanisms that drive skeletal elongation, little is known about how individual bones reach different lengths in any species. Here, we directly compare the transcriptomes of homologous growth cartilages of the mouse (Mus musculus) and bipedal jerboa (Jaculus jaculus), the latter of which has "mouse-like" arms but extremely long metatarsals of the feet. Intersecting gene-expression differences in metatarsals and forearms of the two species revealed that about 10% of orthologous genes are associated with the disproportionately rapid elongation of neonatal jerboa feet. These include genes and enriched pathways not previously associated with endochondral elongation as well as those that might diversify skeletal proportion in addition to their known requirements for bone growth throughout the skeleton. We also identified transcription regulators that might act as "nodes" for sweeping differences in genome expression between species. Among these, Shox2, which is necessary for proximal limb elongation, has gained expression in jerboa metatarsals where it has not been detected in other vertebrates. We show that Shox2 is sufficient to increase mouse distal limb length, and a nearby putative cis-regulatory region is preferentially accessible in jerboa metatarsals. In addition to mechanisms that might directly promote growth, we found evidence that jerboa foot elongation may occur in part by de-repressing latent growth potential. The genes and pathways that we identified here provide a framework to understand the modular genetic control of skeletal growth and the remarkable malleability of vertebrate limb proportion.
Keywords: evolution of development; limb development; skeletal growth; skeletal proportion.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Cellular and molecular mechanisms that shape the development and evolution of tail vertebral proportion in mice and jerboas.bioRxiv [Preprint]. 2024 Oct 26:2024.10.25.620311. doi: 10.1101/2024.10.25.620311. bioRxiv. 2024. PMID: 39484405 Free PMC article. Preprint.
-
The lesser Egyptian jerboa, Jaculus jaculus: a unique rodent model for evolution and development.Cold Spring Harb Protoc. 2011 Dec 1;2011(12):1451-6. doi: 10.1101/pdb.emo066704. Cold Spring Harb Protoc. 2011. PMID: 22135653
-
Metatarsal fusion resisted bending as jerboas (Dipodidae) transitioned from quadrupedal to bipedal.Proc Biol Sci. 2022 Oct 12;289(1984):20221322. doi: 10.1098/rspb.2022.1322. Epub 2022 Oct 5. Proc Biol Sci. 2022. PMID: 36196542 Free PMC article.
-
Developmental and Evolutionary Allometry of the Mammalian Limb Skeleton.Integr Comp Biol. 2019 Nov 1;59(5):1356-1368. doi: 10.1093/icb/icz082. Integr Comp Biol. 2019. PMID: 31180500 Free PMC article. Review.
-
Hox D genes and the fin-to-limb transition: Insights from fish studies.Genesis. 2018 Jan;56(1). doi: 10.1002/dvg.23069. Epub 2017 Oct 13. Genesis. 2018. PMID: 28913932 Review.
Cited by
-
Cellular and molecular mechanisms that shape the development and evolution of tail vertebral proportion in mice and jerboas.bioRxiv [Preprint]. 2024 Oct 26:2024.10.25.620311. doi: 10.1101/2024.10.25.620311. bioRxiv. 2024. PMID: 39484405 Free PMC article. Preprint.
-
The genetic architecture of the human skeletal form.bioRxiv [Preprint]. 2023 Jan 3:2023.01.03.521284. doi: 10.1101/2023.01.03.521284. bioRxiv. 2023. Update in: Science. 2023 Jul 21;381(6655):eadf8009. doi: 10.1126/science.adf8009. PMID: 36712136 Free PMC article. Updated. Preprint.
-
Adapting to change: insights from new organisms in cell and developmental biology.Development. 2024 Oct 15;151(20):dev204389. doi: 10.1242/dev.204389. Epub 2024 Oct 14. Development. 2024. PMID: 39399900
-
The genetic architecture and evolution of the human skeletal form.Science. 2023 Jul 21;381(6655):eadf8009. doi: 10.1126/science.adf8009. Epub 2023 Jul 21. Science. 2023. PMID: 37471560 Free PMC article.
-
Comparative transcriptomics in serial organs uncovers early and pan-organ developmental changes associated with organ-specific morphological adaptation.Nat Commun. 2025 Jan 17;16(1):768. doi: 10.1038/s41467-025-55826-w. Nat Commun. 2025. PMID: 39824799 Free PMC article.
References
-
- Kronenberg HM (2003). Developmental regulation of the growth plate. Nature 423, 332–336. - PubMed
-
- Krakow D, and Rimoin DL (2010). The skeletal dysplasias. Genet. Med 12, 327–341. - PubMed
-
- Christians JK, Bingham VK, Oliver FK, Heath TT, and Keightley PD (2003). Characterization of a QTL affecting skeletal size in mice. Mamm. Genome 14, 175–183. - PubMed
