

Phosphatidylserine receptors enhance SARS-CoV-2 infection
- PMID: 34797899
- PMCID: PMC8641883
- DOI: 10.1371/journal.ppat.1009743
Phosphatidylserine receptors enhance SARS-CoV-2 infection
Abstract
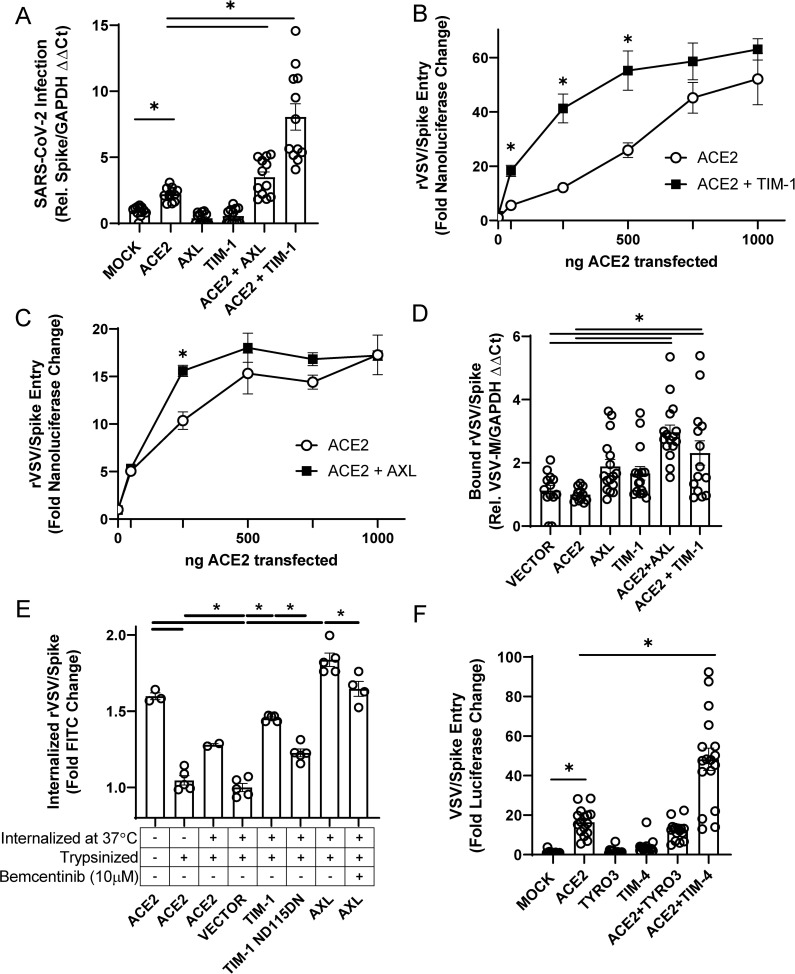
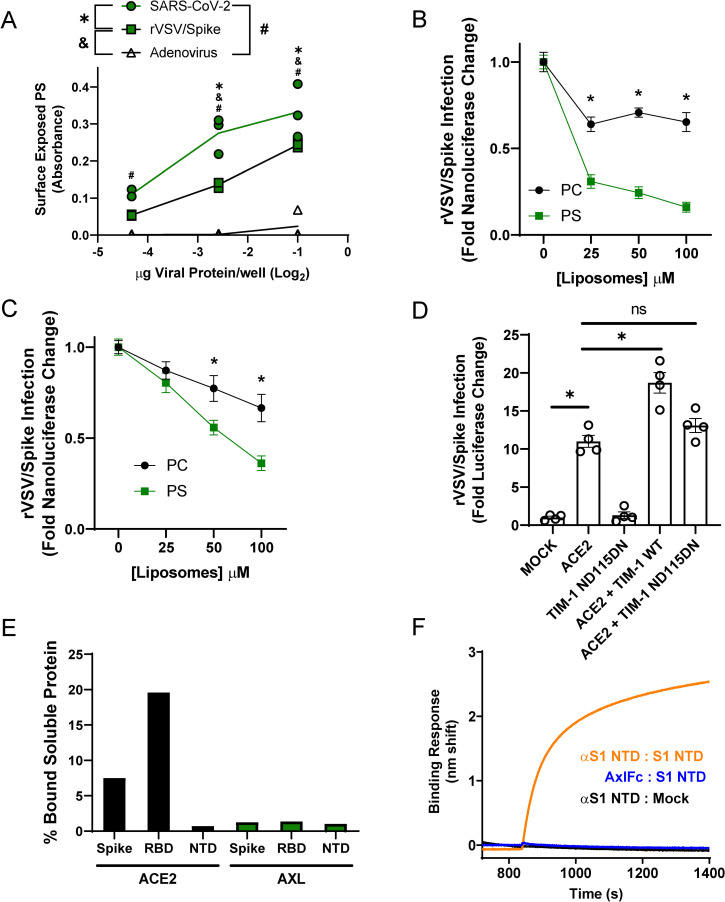
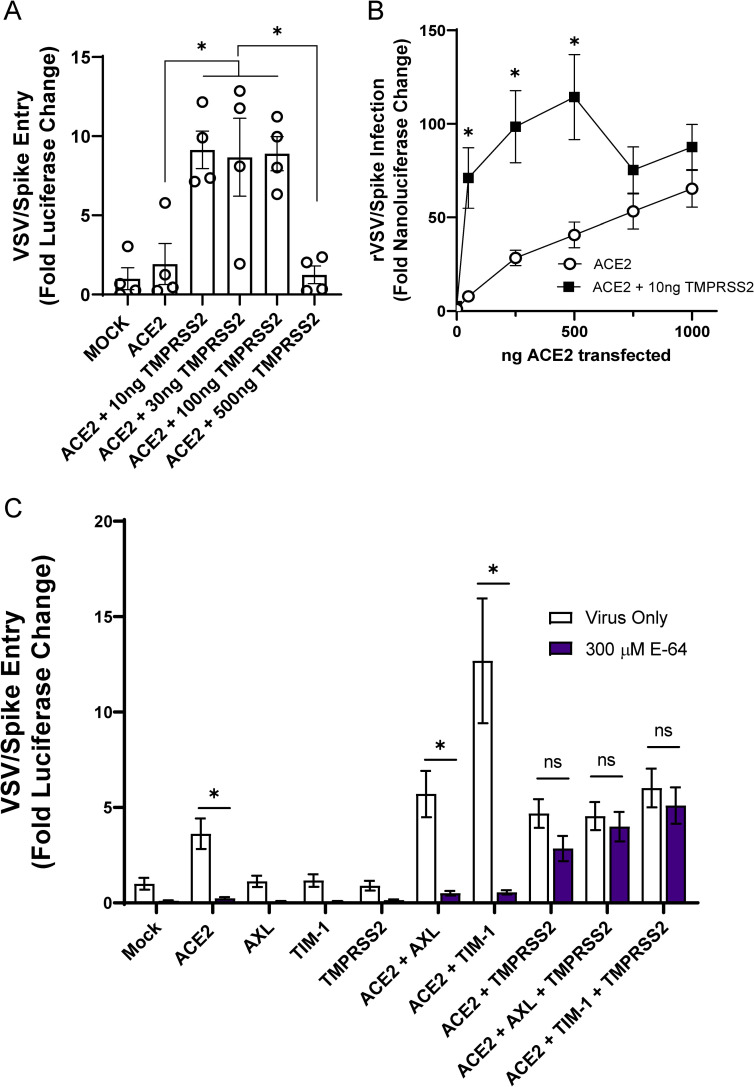
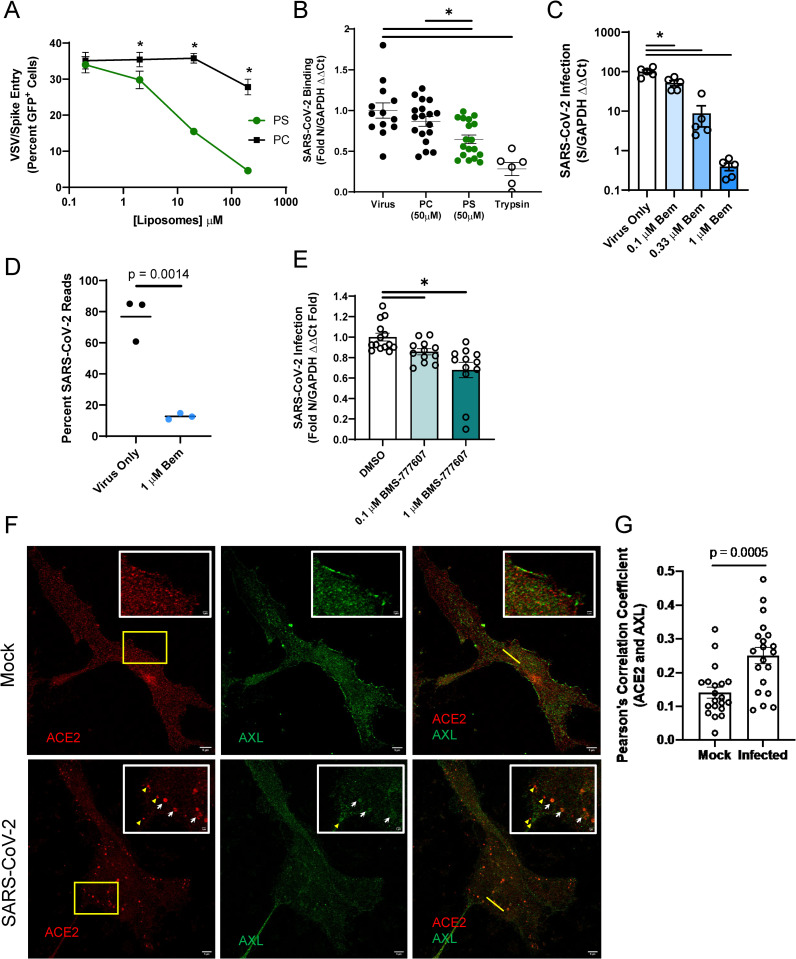
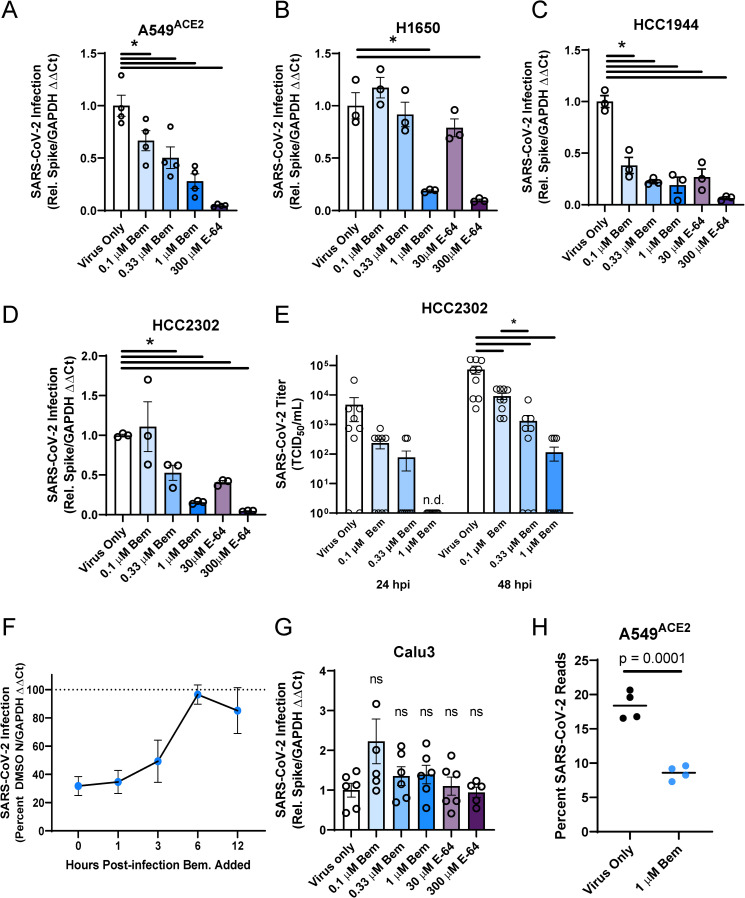
Phosphatidylserine (PS) receptors enhance infection of many enveloped viruses through virion-associated PS binding that is termed apoptotic mimicry. Here we show that this broadly shared uptake mechanism is utilized by SARS-CoV-2 in cells that express low surface levels of ACE2. Expression of members of the TIM (TIM-1 and TIM-4) and TAM (AXL) families of PS receptors enhance SARS-CoV-2 binding to cells, facilitate internalization of fluorescently-labeled virions and increase ACE2-dependent infection of SARS-CoV-2; however, PS receptors alone did not mediate infection. We were unable to detect direct interactions of the PS receptor AXL with purified SARS-CoV-2 spike, contrary to a previous report. Instead, our studies indicate that the PS receptors interact with PS on the surface of SARS-CoV-2 virions. In support of this, we demonstrate that: 1) significant quantities of PS are located on the outer leaflet of SARS-CoV-2 virions, 2) PS liposomes, but not phosphatidylcholine liposomes, reduced entry of VSV/Spike pseudovirions and 3) an established mutant of TIM-1 which does not bind to PS is unable to facilitate entry of SARS-CoV-2. As AXL is an abundant PS receptor on a number of airway lines, we evaluated small molecule inhibitors of AXL signaling such as bemcentinib for their ability to inhibit SARS-CoV-2 infection. Bemcentinib robustly inhibited virus infection of Vero E6 cells as well as multiple human lung cell lines that expressed AXL. This inhibition correlated well with inhibitors that block endosomal acidification and cathepsin activity, consistent with AXL-mediated uptake of SARS-CoV-2 into the endosomal compartment. We extended our observations to the related betacoronavirus mouse hepatitis virus (MHV), showing that inhibition or ablation of AXL reduces MHV infection of murine cells. In total, our findings provide evidence that PS receptors facilitate infection of the pandemic coronavirus SARS-CoV-2 and suggest that inhibition of the PS receptor AXL has therapeutic potential against SARS-CoV-2.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: GG, DM, and EC are employees of BerGenBio ASA, a company with financial interests in this field. JBL is a former employee of BerGenBio ASA. Partial funding was provided by BerGenBio ASA. JM receives licensing royalties from the NIH and UTSW for distribution of human tumor lines. No other authors have competing interests to declare.
Figures

Update of
-
Phosphatidylserine Receptors Enhance SARS-CoV-2 Infection: AXL as a Therapeutic Target for COVID-19.bioRxiv [Preprint]. 2021 Jun 24:2021.06.15.448419. doi: 10.1101/2021.06.15.448419. bioRxiv. 2021. Update in: PLoS Pathog. 2021 Nov 19;17(11):e1009743. doi: 10.1371/journal.ppat.1009743. PMID: 34159331 Free PMC article. Updated. Preprint.
References
-
- Wrobel AG, Benton DJ, Xu P, Roustan C, Martin SR, Rosenthal PB, et al. Author Correction: SARS-CoV-2 and bat RaTG13 spike glycoprotein structures inform on virus evolution and furin-cleavage effects. Nat Struct Mol Biol. 2020;27(10):1001. Epub 2020/08/28. doi: 10.1038/s41594-020-0509-2 ; PubMed Central PMCID: PMC7448700. - DOI - PMC - PubMed
-
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020;181(2):271–80.e8. Epub 2020/03/05. doi: 10.1016/j.cell.2020.02.052 ; PubMed Central PMCID: PMC7102627. - DOI - PMC - PubMed
-
- Coutard B, Valle C, de Lamballerie X, Canard B, Seidah NG, Decroly E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res. 2020;176:104742. Epub 2020/02/15. doi: 10.1016/j.antiviral.2020.104742 ; PubMed Central PMCID: PMC7114094. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
